Klasyfikacja bakterii ze względu na ich środowisko. Zasady klasyfikacji mikroorganizmów
2.1. Systematyka i nazewnictwo drobnoustrojów
Świat drobnoustrojów można podzielić na formy komórkowe i niekomórkowe. Komórkowe formy drobnoustrojów reprezentowane są przez bakterie, grzyby i pierwotniaki. Można je nazwać mikroorganizmami. Formy niekomórkowe są reprezentowane przez wirusy, wiroidy i priony.
Nowa klasyfikacja drobnoustrojów komórkowych obejmuje następujące jednostki taksonomiczne: domeny, królestwa, typy, klasy, rzędy, rodziny, rodzaje, gatunki. Klasyfikacja mikroorganizmów opiera się na ich pokrewieństwie genetycznym, a także właściwościach morfologicznych, fizjologicznych, antygenowych i biologii molekularnej.
Wirusy są często uważane nie za organizmy, ale za autonomiczne struktury genetyczne, dlatego będą rozpatrywane osobno.
Formy komórkowe drobnoustrojów dzielą się na trzy domeny. Domeny Bakteria I Archebakterie obejmują drobnoustroje o strukturze komórkowej typu prokariotycznego. Przedstawiciele domen Eukarya są eukariontami. Składa się z 4 królestw:
Królestwa grzybów (Grzyby, Eumycota);
królestwa pierwotniaków (pierwotniaki);
królestwa Chromista(płyty chromowane);
Drobnoustroje o nieokreślonej pozycji taksonomicznej (Mikrospora, mikrosporydia).
Różnice w organizacji komórek prokariotycznych i eukariotycznych przedstawiono w tabeli. 2.1.
Tabela 2.1. Znaki komórki prokariotycznej i eukariotycznej
2.2. Klasyfikacja i morfologia bakterii
Od tego słowa pochodzi termin „bakterie”. bakteria, co to znaczy kij? Bakterie są prokariotami. Dzielą się na dwie domeny: Bakteria I Archebakterie. Bakterie zawarte w domenie Archebakterie, reprezentują jedną z najstarszych form życia. Mają cechy strukturalne ściany komórkowej (brak peptydoglikanu) i rybosomalnego RNA. Nie ma wśród nich patogenów chorób zakaźnych.
W ramach domeny bakterie dzieli się na następujące kategorie taksonomiczne: klasa, typ, rząd, rodzina, rodzaj, gatunek. Jedną z głównych kategorii taksonomicznych jest gatunek. Gatunek to zbiór osobników o tym samym pochodzeniu i genotypie, połączonych podobnymi cechami, które odróżniają je od innych przedstawicieli rodzaju. Nazwa gatunkowa odpowiada nomenklaturze binarnej, tj. składa się z dwóch słów. Na przykład czynnik wywołujący błonicę jest zapisywany jako Corynebacterium diphtheriae. Pierwsze słowo jest nazwą rodzaju i jest pisane wielką literą, drugie słowo oznacza gatunek i jest pisane małą literą.
W przypadku ponownej wzmianki o gatunku nazwę rodzajową skraca się do początkowej litery, np. C. błonica.
Nazywa się zbiór jednorodnych mikroorganizmów wyizolowanych na pożywce, charakteryzujących się podobnymi właściwościami morfologicznymi, barwnymi (w stosunku do barwników), kulturowymi, biochemicznymi i antygenowymi czysta kultura. Nazywa się czystą kulturą mikroorganizmów wyizolowanych z określonego źródła i różniących się od innych przedstawicieli gatunku napięcie. Bliskie pojęciu „szczepu” jest pojęcie „klonu”. Klon to zbiór potomków wyhodowanych z pojedynczej komórki drobnoustroju.
Aby oznaczyć pewne zbiory mikroorganizmów, które różnią się pewnymi właściwościami, stosuje się przyrostek „var” (odmiana), dlatego mikroorganizmy, w zależności od charakteru różnic, określa się jako morfowary (różnica w morfologii), produkty oporne (różnica w oporności na przykład na antybiotyki), serotypy (różnica w antygenach), fagowary (różnica we wrażliwości na bakteriofagi), biowary (różnica we właściwościach biologicznych), chemowary (różnica we właściwościach biochemicznych) itp.
Wcześniej podstawą klasyfikacji bakterii była cecha strukturalna ściany komórkowej. Podział bakterii ze względu na cechy strukturalne ściany komórkowej wiąże się z możliwą zmiennością ich zabarwienia na ten lub inny kolor metodą Grama. Według tej metody, zaproponowanej w 1884 roku przez duńskiego naukowca H. Grama, w zależności od wyniku barwienia, bakterie dzieli się na Gram-dodatnie, barwione na niebiesko-fioletowo i Gram-ujemne, barwione na czerwono.
Obecnie klasyfikacja opiera się na stopniu pokrewieństwa genetycznego, w oparciu o badanie struktury genomu rybosomalnego RNA (rRNA) (patrz rozdz. 5), określenie procentowego udziału par guaniny-cytozyny (par GC) w genomie, konstruowanie mapę restrykcyjną genomu i badanie stopnia hybrydyzacji. Uwzględnia się także wskaźniki fenotypowe: stosunek do barwienia metodą Grama, właściwości morfologiczne, kulturowe i biochemiczne, strukturę antygenową.
Domena Bakteria obejmuje 23 typy, z których następujące mają znaczenie medyczne.
Większość bakterii Gram-ujemnych grupuje się w gromadę Proteobakterie(nazwany na cześć greckiego boga Odmieniec, mogących przybierać różne formy). Typ Proteobakterie podzielony na 5 klas:
Klasa Alfaproteobakterie(narodziny Rickettsia, Orientia, Erlichia, Bartonella, Brucella);
Klasa Betaproteobakterie(narodziny Bordetella, Burholderia, Neisseria, Spirillum);
Klasa Gammaproteobakterie(przedstawiciele rodziny Enterobakterie poród Francisella, Legionella, Coxiella, Pseudomonas, Vibrio);
Klasa Deltaproteobakterie(rodzaj Bilofilia);
Klasa Epsilonproteobakterie(narodziny Campilobacter, Helicobacter). Bakterie Gram-ujemne obejmują również następujące typy:
typ Chlamydie(narodziny Chlamydia, Chlamydophila), typ Krętki(narodziny Spirocheta, Borrelia, Treponema, Leptospira); typ Bacteroides(narodziny Bacteroides, Prevotella, Porphyromonas).
Bakterie Gram-dodatnie dzielą się na następujące typy:
Typ Firmicutes zawiera klasę Clostridium(narodziny Clostridium, Peptococcus), Klasa Pałeczki (Listeria, Staphylococcus, Lactobacillus, Streptococcus) i klasa Mollicutes(narodziny mykoplazma, ureaplazma), które są bakteriami nieposiadającymi ściany komórkowej;
typ Actinobakterie(narodziny Actinomyces, Micrococcus, Corynebacterium, Mycobacterium, Gardnerella, Bifidobacterium, Propionibacterium, Mobiluncus).
2.2.1. Formy morfologiczne bakterii
Istnieje kilka głównych form bakterii: kokosowata, w kształcie pręcika, zwinięta i rozgałęziona (ryc. 2.1).
Formy kuliste lub ziarniaki- bakterie kuliste o wielkości 0,5-1 mikrona, które zgodnie z ich względnym położeniem dzielą się na mikrokoki, diplokoki, paciorkowce, tetrakoki, sarcina i gronkowce.
Micrococci (z greckiego. mikro- mały) - oddzielnie zlokalizowane komórki.
Diplococci (z greckiego. dyplomaci- podwójne) lub sparowane ziarniaki znajdują się w parach (pneumokoki, gonokoki, meningokoki), ponieważ komórki nie rozdzielają się po podziale. Pneumokoki (czynnik wywołujący zapalenie płuc) mają lancetowaty kształt po przeciwnych stronach, a rzeżączka (czynnik wywołujący rzeżączkę) i meningokok (czynnik wywołujący rzeżączkę)
 Ryż. 2.1. Kształty bakterii
Ryż. 2.1. Kształty bakterii
czynnik wywołujący epidemiczne zapalenie opon mózgowo-rdzeniowych) mają kształt ziaren kawy, których wklęsła powierzchnia jest zwrócona ku sobie.
Streptococci (z greckiego. streptos- łańcuch) - komórki o okrągłym lub wydłużonym kształcie, tworzące łańcuch w wyniku podziału komórek w tej samej płaszczyźnie i utrzymania połączenia między nimi w miejscu podziału.
Sarcyny (od łac. sarcyna- pęczek, bela) są ułożone w postaci pakietów po 8 ziarniaków lub więcej, ponieważ powstają podczas podziału komórek w trzech wzajemnie prostopadłych płaszczyznach.
Staphylococcus (z greckiego. staphyl- kiść winogron) - ziarniaki powstałe w postaci kiści winogron w wyniku podziału w różnych płaszczyznach.
Bakterie w kształcie pręta różnią się rozmiarem, kształtem końców komórek i względnym położeniem komórek. Długość komórki wynosi 1-10 µm, grubość 0,5-2 µm. Kije mogą mieć rację
(Escherichia coli itp.) i nieregularny kształt maczugi (Corynebacteria itp.). Do najmniejszych bakterii w kształcie pręcików zalicza się riketsje.
Końce pręcików można odciąć (bacillus wąglika), zaokrąglić (Escherichia coli), zaostrzyć (fusobakterie) lub w formie zgrubienia. W tym drugim przypadku pręt wygląda jak maczuga (Corynebacterium diphtheria).
Lekko zakrzywione pręty nazywane są vibrios (Vibrio cholerae). Większość bakterii w kształcie pałeczek jest rozmieszczonych losowo, ponieważ komórki po podziale oddalają się od siebie. Jeżeli po podziale komórki pozostają połączone wspólnymi fragmentami ściany komórkowej i nie rozchodzą się, wówczas układają się względem siebie pod kątem (Corynebacterium diphtheria) lub tworzą łańcuch (bacillus wąglika).
Skręcone Kształty- bakterie spiralne, które występują w dwóch rodzajach: spirilla i krętki. Spirilla ma wygląd skręconych komórek w kształcie korkociągu z dużymi lokami. Do patogennych spirilli zalicza się czynnik sprawczy sodoku (choroba ukąszenia szczura), a także Campylobacter i Helicobacter, które mają krzywizny przypominające skrzydła latającej mewy. Krętki to cienkie, długie i zawiłe bakterie, które różnią się od spirilli mniejszymi lokami i wzorcem ruchu. Poniżej opisano cechy charakterystyczne ich struktury.
Rozgałęzienie - bakterie w kształcie pręcika, które mogą mieć rozgałęzienia w kształcie litery Y występujące u bifidobakterii, mogą również występować w postaci rozgałęzionych komórek nitkowatych, które mogą się przeplatać, tworząc grzybnię, co obserwuje się u promieniowców.
2.2.2. Struktura komórki bakteryjnej
Strukturę bakterii dobrze zbadano za pomocą mikroskopii elektronowej całych komórek i ich ultracienkich skrawków, a także innymi metodami. Komórka bakteryjna jest otoczona błoną składającą się ze ściany komórkowej i błony cytoplazmatycznej. Pod skorupą znajduje się protoplazma, składająca się z cytoplazmy z wtrąceniami i dziedzicznego aparatu - analogu jądra, zwanego nukleoidem (ryc. 2.2). Istnieją dodatkowe struktury: kapsułka, mikrokapsułka, śluz, wici, pilusy. Niektóre bakterie są zdolne do tworzenia zarodników w niesprzyjających warunkach.
 Ryż. 2.2. Struktura komórki bakteryjnej: 1 - kapsułka; 2 - ściana komórkowa; 3 - błona cytoplazmatyczna; 4 - mezosomy; 5 - nukleoid; 6 - plazmid; 7 - rybosomy; 8 - wtrącenia; 9 - wici; 10 - pili (kosmki)
Ryż. 2.2. Struktura komórki bakteryjnej: 1 - kapsułka; 2 - ściana komórkowa; 3 - błona cytoplazmatyczna; 4 - mezosomy; 5 - nukleoid; 6 - plazmid; 7 - rybosomy; 8 - wtrącenia; 9 - wici; 10 - pili (kosmki)
Ściana komórkowa- mocna, elastyczna struktura, która nadaje bakterii określony kształt i wraz z leżącą pod nią błoną cytoplazmatyczną powstrzymuje wysokie ciśnienie osmotyczne w komórce bakteryjnej. Bierze udział w procesie podziału komórek i transporcie metabolitów, posiada receptory dla bakteriofagów, bakteriocyn i różnych substancji. Najgrubszą ścianę komórkową występują u bakterii Gram-dodatnich (ryc. 2.3). Tak więc, jeśli grubość ściany komórkowej bakterii Gram-ujemnych wynosi około 15-20 nm, wówczas u bakterii Gram-dodatnich może osiągnąć 50 nm lub więcej.
Podstawą ściany komórkowej bakterii jest peptydoglikan. Peptydoglikan jest polimerem. Jest reprezentowany przez równoległe polisacharydowe łańcuchy glikanów składające się z powtarzających się reszt N-acetyloglukozaminy i kwasu N-acetylomuraminowego, połączonych wiązaniem glikozydowym. Wiązanie to zostaje rozerwane przez lizozym, będący acetylmuramidazą.
Tetrapeptyd jest przyłączony do kwasu N-acetylomuraminowego wiązaniami kowalencyjnymi. Tetrapeptyd składa się z L-alaniny, która jest połączona z kwasem N-acetylomuraminowym; D-glutamina, która u bakterii Gram-dodatnich łączy się z L-lizyną, a u bakterii Gram-tri-
 Ryż. 2.3. Schemat architektury ściany komórkowej bakterii
Ryż. 2.3. Schemat architektury ściany komórkowej bakterii
pożyteczne bakterie – z kwasem diaminopimelinowym (DAP), który jest prekursorem lizyny w procesie bakteryjnej biosyntezy aminokwasów i jest unikalnym związkiem występującym wyłącznie w bakteriach; Czwartym aminokwasem jest D-alanina (ryc. 2.4).
Ściana komórkowa bakterii Gram-dodatnich zawiera niewielkie ilości polisacharydów, lipidów i białek. Głównym składnikiem ściany komórkowej tych bakterii jest wielowarstwowy peptydoglikan (mureina, mukopeptyd), stanowiący 40-90% masy ściany komórkowej. Tetrapeptydy różnych warstw peptydoglikanu u bakterii Gram-dodatnich są połączone ze sobą łańcuchami polipeptydowymi złożonymi z 5 reszt glicyny (pentaglicyny), co nadaje peptydoglikanowi sztywną strukturę geometryczną (ryc. 2.4, b). Kowalencyjnie związany z peptydoglikanem ściany komórkowej bakterii Gram-dodatnich kwasy teichojowe(z greckiego techos- ściana), których cząsteczki są łańcuchami 8-50 reszt glicerolu i rybitolu połączonych mostkami fosforanowymi. Kształt i siłę bakterii nadaje sztywna włóknista struktura wielowarstwowego peptydoglikanu z usieciowanymi wiązaniami peptydów.
 Ryż. 2.4. Struktura peptydoglikanu: a - bakterie Gram-ujemne; b - bakterie Gram-dodatnie
Ryż. 2.4. Struktura peptydoglikanu: a - bakterie Gram-ujemne; b - bakterie Gram-dodatnie
Zdolność bakterii Gram-dodatnich do zatrzymywania fioletu goryczki w połączeniu z jodem podczas barwienia metodą Grama (niebiesko-fioletowy kolor bakterii) jest związana z właściwością wielowarstwowego peptydoglikanu do interakcji z barwnikiem. Ponadto późniejsze traktowanie rozmazu bakteryjnego alkoholem powoduje zwężenie porów w peptydoglikanie, a tym samym zatrzymuje barwnik w ścianie komórkowej.
Bakterie Gram-ujemne tracą barwnik pod wpływem alkoholu, co wynika z mniejszej ilości peptydoglikanu (5-10% masy ściany komórkowej); odbarwiają się pod wpływem alkoholu, a pod wpływem fuksyny lub safraniny stają się czerwone. Wynika to z cech strukturalnych ściany komórkowej. Peptydoglikan w ścianie komórkowej bakterii Gram-ujemnych jest reprezentowany przez 1-2 warstwy. Tetrapeptydy warstw są połączone ze sobą bezpośrednim wiązaniem peptydowym pomiędzy grupą aminową DAP jednego tetrapeptydu a grupą karboksylową D-alaniny tetrapeptydu innej warstwy (ryc. 2.4, a). Na zewnątrz peptydoglikanu znajduje się warstwa lipoproteina, połączony z peptydoglikanem poprzez DAP. Śledzony przez zewnętrzna męmbranaŚciana komórkowa.
Zewnętrzna męmbrana jest strukturą mozaikową złożoną z lipopolisacharydów (LPS), fosfolipidów i białek. Jego wewnętrzna warstwa jest reprezentowana przez fosfolipidy, a warstwa zewnętrzna zawiera LPS (ryc. 2.5). Zatem pamięć zewnętrzna
 Ryż. 2.5. Struktura lipopolisacharydu
Ryż. 2.5. Struktura lipopolisacharydu
Brana jest asymetryczna. Błona zewnętrzna LPS składa się z trzech fragmentów:
Lipid A ma konserwatywną strukturę, prawie taką samą jak u bakterii Gram-ujemnych. Lipid A składa się z fosforylowanych jednostek disacharydowych glukozaminy, do których przyłączone są długie łańcuchy kwasów tłuszczowych (patrz ryc. 2.5);
Rdzeń lub rdzeń, część skorupy ziemskiej (od łac. rdzeń- rdzeń), stosunkowo konserwatywna struktura oligosacharydowa;
Wysoce zmienny łańcuch polisacharydowy specyficzny dla O, utworzony przez powtarzanie identycznych sekwencji oligosacharydowych.
LPS jest zakotwiczony w błonie zewnętrznej przez lipid A, który powoduje toksyczność LPS i dlatego jest utożsamiany z endotoksyną. Niszczenie bakterii przez antybiotyki prowadzi do uwolnienia dużych ilości endotoksyn, które mogą wywołać u pacjenta wstrząs endotoksyczny. Rdzeń lub część rdzeniowa LPS rozciąga się od lipidu A. Najbardziej stałą częścią rdzenia LPS jest kwas ketodeoksyoktonowy. O-specyficzny łańcuch polisacharydowy rozciągający się od rdzenia cząsteczki LPS,
składający się z powtarzających się jednostek oligosacharydowych, określa serogrupę, serotyp (rodzaj bakterii wykrywany za pomocą surowicy odpornościowej) konkretnego szczepu bakterii. Zatem koncepcja LPS jest powiązana z koncepcją antygenu O, dzięki któremu można różnicować bakterie. Zmiany genetyczne mogą prowadzić do defektów, skrócenia bakteryjnego LPS i w efekcie do pojawienia się szorstkich kolonii form R, które tracą specyficzność antygenu O.
Nie wszystkie bakterie Gram-ujemne mają kompletny łańcuch polisacharydowy specyficzny dla O, składający się z powtarzających się jednostek oligosacharydowych. W szczególności bakterie z rodzaju Neisseria mają krótki glikolipid zwany lipooligosacharydem (LOS). Jest porównywalna z formą R, która utraciła specyficzność antygenu O, obserwowaną u zmutowanych szczepów szorstkich E coli. Struktura LZO przypomina strukturę glikosfingolipidu ludzkiej błony cytoplazmatycznej, zatem LZO naśladuje drobnoustrój, umożliwiając mu uniknięcie odpowiedzi immunologicznej gospodarza.
Białka macierzy błony zewnętrznej przenikają ją w taki sposób, że cząsteczki białka tzw porinami, graniczą z porami hydrofilowymi, przez które przechodzi woda i małe cząsteczki hydrofilowe o masie względnej do 700 D.
Pomiędzy błoną zewnętrzną a cytoplazmatyczną znajduje się Przestrzeń periplazmatyczna, lub peryplazmę zawierającą enzymy (proteazy, lipazy, fosfatazy, nukleazy, β-laktamazy), a także składniki systemów transportowych.
Kiedy synteza ściany komórkowej bakterii zostaje zakłócona pod wpływem lizozymu, penicyliny, czynników ochronnych organizmu i innych związków, powstają komórki o zmienionym (często kulistym) kształcie: protoplasty- bakterie całkowicie pozbawione ściany komórkowej; sferoplasty- bakterie z częściowo zachowaną ścianą komórkową. Po usunięciu inhibitora ściany komórkowej, takie zmienione bakterie mogą ulec odwróceniu, tj. uzyskać pełną ścianę komórkową i przywrócić jej pierwotny kształt.
Bakterie typu sferoidalnego lub protoplastowego, które pod wpływem antybiotyków lub innych czynników utraciły zdolność do syntezy peptydoglikanu i są w stanie się rozmnażać, nazywane są bakteriami Kształty L(od nazwy Instytutu D. Listera, gdzie po raz pierwszy
Zostały zbadane). Formy L mogą również powstawać w wyniku mutacji. Są to wrażliwe osmotycznie, kuliste komórki o kształcie kolby, różnej wielkości, w tym także te przechodzące przez filtry bakteryjne. Niektóre formy L (niestabilne), po usunięciu czynnika wywołującego zmiany w bakteriach, mogą się odwrócić, powracając do pierwotnej komórki bakteryjnej. Formy L mogą być wytwarzane przez wiele patogenów chorób zakaźnych.
Błona cytoplazmatyczna w mikroskopii elektronowej ultracienkich przekrojów jest to membrana trójwarstwowa (2 ciemne warstwy, każda o grubości 2,5 nm, oddzielone jasną warstwą pośrednią). Strukturalnie przypomina plazmalemmę komórek zwierzęcych i składa się z podwójnej warstwy lipidów, głównie fosfolipidów, z osadzoną powierzchnią i integralnymi białkami, które wydają się przenikać przez strukturę błony. Część z nich to permeazy biorące udział w transporcie substancji. W przeciwieństwie do komórek eukariotycznych, błona cytoplazmatyczna komórki bakteryjnej nie zawiera steroli (z wyjątkiem mykoplazm).
Błona cytoplazmatyczna jest dynamiczną strukturą zawierającą ruchome elementy, dlatego uważa się ją za mobilną strukturę płynną. Otacza zewnętrzną część cytoplazmy bakterii i bierze udział w regulacji ciśnienia osmotycznego, transporcie substancji i metabolizmie energetycznym komórki (dzięki enzymom łańcucha transportu elektronów, trifosfatazy adenozyny - ATPazy itp.). Przy nadmiernym wzroście (w porównaniu ze wzrostem ściany komórkowej) błona cytoplazmatyczna tworzy wgłobienia - wgłobienia w postaci kompleksowo skręconych struktur błonowych, zwanych mezosomy. Mniej skomplikowane skręcone struktury nazywane są błonami wewnątrzcytoplazmatycznymi. Rola mezosomów i błon wewnątrzcytoplazmatycznych nie jest w pełni poznana. Sugeruje się nawet, że są one artefaktem powstającym po przygotowaniu (utrwaleniu) próbki do mikroskopii elektronowej. Niemniej jednak uważa się, że pochodne błony cytoplazmatycznej biorą udział w podziale komórek, dostarczając energii do syntezy ściany komórkowej oraz biorą udział w wydzielaniu substancji, sporulacji, tj. w procesach o dużym zużyciu energii. Cytoplazma zajmuje główną objętość bakterii
komórka i składa się z rozpuszczalnych białek, kwasów rybonukleinowych, inkluzji i licznych małych granulek - rybosomów, odpowiedzialnych za syntezę (translację) białek.
Rybosomy bakterie mają wielkość około 20 nm i współczynnik sedymentacji 70S, w przeciwieństwie do rybosomów 80S charakterystycznych dla komórek eukariotycznych. Dlatego niektóre antybiotyki, wiążąc się z rybosomami bakteryjnymi, hamują syntezę białek bakteryjnych, nie wpływając na syntezę białek w komórkach eukariotycznych. Rybosomy bakteryjne mogą dysocjować na dwie podjednostki: 50S i 30S. rRNA jest konserwatywnym elementem bakterii („molekularnym zegarem” ewolucji). 16S rRNA jest częścią małej podjednostki rybosomu, a 23S rRNA jest częścią dużej podjednostki rybosomu. Badanie 16S rRNA jest podstawą systematyki genów, pozwalającą ocenić stopień pokrewieństwa organizmów.
Cytoplazma zawiera różne wtrącenia w postaci granulek glikogenu, polisacharydów, kwasu β-hydroksymasłowego i polifosforanów (wolutyny). Kumulują się w przypadku nadmiaru składników odżywczych w środowisku i pełnią funkcję substancji rezerwowych na potrzeby żywieniowe i energetyczne.
Wolutin ma powinowactwo do barwników zasadowych i można go łatwo wykryć specjalnymi metodami barwienia (na przykład według Neissera) w postaci granulek metachromatycznych. W przypadku błękitu toluidynowego lub błękitu metylenowego wolutyna jest zabarwiona na czerwono-fioletowo, a cytoplazma bakterii jest zabarwiona na niebiesko. Charakterystyczny układ ziaren wolutyny u prątka błonicy ujawnia się w postaci intensywnie wybarwionych biegunów komórkowych. Metachromatyczne zabarwienie wolutyny wiąże się z dużą zawartością spolimeryzowanego nieorganicznego polifosforanu. Pod mikroskopem elektronowym wyglądają jak granulki o dużej gęstości elektronowej i wielkości 0,1–1 mikrona.
Nukleoid- odpowiednik jądra u bakterii. Znajduje się w centralnej strefie bakterii w postaci dwuniciowego DNA, ciasno upakowanego jak kula. Nukleoid bakterii, w przeciwieństwie do eukariontów, nie ma otoczki jądrowej, jąderka i podstawowych białek (histonów). Większość bakterii zawiera jeden chromosom, reprezentowany przez cząsteczkę DNA zamkniętą w pierścieniu. Ale niektóre bakterie mają dwa chromosomy w kształcie pierścienia (V. cholerae) i chromosomy liniowe (patrz sekcja 5.1.1). Nukleoid ujawnia się w mikroskopie świetlnym po zabarwieniu barwnikami specyficznymi dla DNA
metody: według Feulgena lub według Romanovsky-Giemsa. Na obrazach dyfrakcji elektronów ultracienkich skrawków bakterii nukleoid pojawia się jako jasne strefy z włóknistymi, nitkowatymi strukturami DNA związanymi w pewnych obszarach z błoną cytoplazmatyczną lub mezosomem zaangażowanym w replikację chromosomu.
Oprócz nukleoidu komórka bakteryjna zawiera pozachromosomalne czynniki dziedziczności - plazmidy (patrz sekcja 5.1.2), które są kowalencyjnie zamkniętymi pierścieniami DNA.
Kapsułka, mikrokapsułka, śluz.Kapsuła - struktura śluzowa o grubości ponad 0,2 mikrona, mocno związana ze ścianą komórkową bakterii i posiadająca wyraźnie określone granice zewnętrzne. Kapsułka jest widoczna w rozmazach odciskowych materiału patologicznego. W czystych kulturach bakteryjnych kapsułka tworzy się rzadziej. Wykrywa się go za pomocą specjalnych metod barwienia rozmazu według Burri-Ginsa, co tworzy negatywny kontrast substancji kapsułki: atrament tworzy ciemne tło wokół kapsułki. Kapsułka składa się z polisacharydów (egzopolisacharydów), czasami z polipeptydów, na przykład w prątku wąglika składa się z polimerów kwasu D-glutaminowego. Kapsułka jest hydrofilowa i zawiera dużą ilość wody. Zapobiega fagocytozie bakterii. Kapsułka jest antygenowa: przeciwciała skierowane przeciwko kapsułce powodują jej powiększenie (reakcja na obrzęk kapsułki).
Tworzy się wiele bakterii mikrokapsułka- tworzenie się śluzu o grubości mniejszej niż 0,2 mikrona, wykrywalnego jedynie za pomocą mikroskopii elektronowej.
Należy go odróżnić od kapsułki szlam -śluzowate egzopolisacharydy, które nie mają wyraźnych granic zewnętrznych. Śluz jest rozpuszczalny w wodzie.
Śluzowe egzopolisacharydy są charakterystyczne dla śluzowatych szczepów Pseudomonas aeruginosa, często spotykanych w plwocinie pacjentów z mukowiscydozą. Egzopolisacharydy bakteryjne biorą udział w adhezji (przyklejaniu się do podłoża); nazywane są również glikokaliksem.
Kapsułka i śluz chronią bakterie przed uszkodzeniem i wysychaniem, ponieważ będąc hydrofilowymi, dobrze wiążą wodę i uniemożliwiają działanie czynników ochronnych makroorganizmu i bakteriofagów.
Wici bakterie determinują ruchliwość komórki bakteryjnej. Wici to cienkie włókna, które się przyjmują
Pochodzą z błony cytoplazmatycznej i są dłuższe niż sama komórka. Grubość wici wynosi 12-20 nm, długość 3-15 µm. Składają się z trzech części: spiralnego włókna, haczyka i korpusu podstawowego zawierającego pręt ze specjalnymi krążkami (jedna para krążków u bakterii Gram-dodatnich i dwie pary krążków u bakterii Gram-ujemnych). Wici są przymocowane do błony cytoplazmatycznej i ściany komórkowej za pomocą krążków. Stwarza to efekt silnika elektrycznego z prętem - wirnikiem - obracającym wici. Jako źródło energii wykorzystywana jest różnica potencjałów protonów na błonie cytoplazmatycznej. Mechanizm rotacji zapewnia syntetaza protonowa ATP. Prędkość obrotowa wici może osiągnąć 100 obr./s. Jeśli bakteria ma kilka wici, zaczynają się one obracać synchronicznie, splatając się w jeden pakiet, tworząc rodzaj śmigła.
Wici są zbudowane z białka zwanego flageliną. (rozłóg- wić), która jest antygenem - tzw. antygenem H. Podjednostki flageliny są skręcone spiralnie.
Liczba wici u różnych gatunków bakterii waha się od jednej (monotrichus) u Vibrio cholerae do dziesiątek i setek rozciągających się wzdłuż obwodu bakterii (peritrichus), u Escherichia coli, Proteus itp. Lofotrichy mają wiązkę wici na jednym końcu komórki. Amphitrichy ma jedną wici lub wiązkę wici na przeciwległych końcach komórki.
Wici wykrywa się za pomocą mikroskopii elektronowej preparatów pokrytych metalami ciężkimi lub w mikroskopie świetlnym po obróbce specjalnymi metodami polegającymi na wytrawianiu i adsorpcji różnych substancji prowadzących do zwiększenia grubości wici (na przykład po srebrzeniu).
Kosmki lub pilusy (fimbrie)- formacje nitkowate, cieńsze i krótsze (3-10 nm * 0,3-10 µm) niż wici. Pili wystają z powierzchni komórki i składają się z pilinu białkowego. Znanych jest kilka rodzajów pilusów. Pili typu ogólnego odpowiadają za przyczepność do podłoża, odżywianie i metabolizm wody i soli. Jest ich mnóstwo – po kilkaset na komórkę. Pili płciowe (1-3 na komórkę) tworzą kontakt między komórkami, przekazując między sobą informację genetyczną poprzez koniugację (patrz rozdział 5). Szczególnie interesujące są pilusy typu IV, których końcówki są hydrofobowe, w wyniku czego ulegają zwijaniu, zwane też włoskami lokowymi. Lokalizacja
Znajdują się na biegunach komórki. Te pilusy znajdują się w bakteriach chorobotwórczych. Mają właściwości antygenowe, umożliwiają kontakt bakterii z komórką gospodarza i biorą udział w tworzeniu biofilmu (patrz rozdział 3). Wiele pilusów jest receptorami dla bakteriofagów.
Spory - osobliwa forma bakterii spoczynkowych o strukturze ściany komórkowej Gram-dodatniego typu. Bakterie tworzące przetrwalniki z rodzaju Bakcyl, w których wielkość zarodników nie przekracza średnicy komórki, nazywane są prątkami. Bakterie tworzące przetrwalniki, u których wielkość zarodników przekracza średnicę komórki, dlatego przyjmują kształt wrzeciona, nazywane są bakteriami tworzącymi przetrwalniki. Clostridia, na przykład bakterie z rodzaju Clostridium(od łac. Clostridium- wrzeciono). Zarodniki są kwasoodporne, dlatego metodą Aujeszky'ego lub Ziehl-Neelsena wybarwia się je na czerwono, a komórkę wegetatywną na niebiesko.
Sporulacja, kształt i umiejscowienie zarodników w komórce (wegetatywnej) są właściwością gatunkową bakterii, która pozwala je odróżnić. Kształt zarodników może być owalny lub kulisty, lokalizacja w komórce jest końcowa, tj. na końcu pałeczki (w czynniku wywołującym tężec), podkońcowym - bliżej końca pałeczki (w czynnikach powodujących zatrucie jadem kiełbasianym, zgorzel gazową) i centralnym (w prątku wąglika).
Proces sporulacji (sporulacji) przebiega przez szereg etapów, podczas których część cytoplazmy i chromosomu bakteryjnej komórki wegetatywnej zostaje oddzielona, otoczona wrastającą błoną cytoplazmatyczną – powstaje prospora.
Protoplast prospory zawiera nukleoid, system syntezy białek i system wytwarzania energii oparty na glikolizie. Cytochromów nie ma nawet u tlenowców. Nie zawiera ATP, energia do kiełkowania magazynowana jest w postaci fosforanu 3-glicerolu.
Prospor jest otoczony dwiema błonami cytoplazmatycznymi. Nazywa się warstwę otaczającą wewnętrzną błonę zarodnika ściana zarodników, składa się z peptydoglikanu i jest głównym źródłem ściany komórkowej podczas kiełkowania zarodników.
Pomiędzy błoną zewnętrzną a ścianą zarodników tworzy się gruba warstwa składająca się z peptydoglikanu, który ma wiele wiązań poprzecznych - kora.
Znajduje się na zewnątrz zewnętrznej błony cytoplazmatycznej skorupa zarodników, zbudowane z białek keratynowych,
posiadające wiele wewnątrzcząsteczkowych wiązań dwusiarczkowych. Powłoka ta zapewnia odporność na czynniki chemiczne. Zarodniki niektórych bakterii mają dodatkową osłonę - egzosporium charakter lipoproteinowy. W ten sposób tworzy się wielowarstwowa, słabo przepuszczalna otoczka.
Sporulacji towarzyszy intensywne zużycie przez prosporę, a następnie rozwijającą się otoczkę zarodników kwasu dipikolinowego i jonów wapnia. Zarodnik zyskuje odporność na ciepło, co jest związane z obecnością w nim dipikolinianu wapnia.
Zarodnik może przetrwać przez długi czas ze względu na obecność wielowarstwowej otoczki, dipikolinianu wapnia, niską zawartość wody i powolne procesy metaboliczne. Na przykład w glebie patogeny wąglika i tężca mogą przetrwać dziesięciolecia.
W sprzyjających warunkach zarodniki kiełkują, przechodząc przez trzy kolejne etapy: aktywację, inicjację, wzrost. W tym przypadku z jednego zarodnika powstaje jedna bakteria. Aktywacja to gotowość do kiełkowania. W temperaturze 60-80°C zarodniki aktywują się do kiełkowania. Inicjacja kiełkowania trwa kilka minut. Faza wzrostu charakteryzuje się szybkim wzrostem, któremu towarzyszy zniszczenie łupiny i pojawienie się siewki.
2.2.3. Cechy strukturalne krętków, riketsjów, chlamydii, promieniowców i mykoplazm
Krętki- cienkie, długie i zawiłe bakterie. Składają się z zewnętrznej błoniastej ściany komórkowej otaczającej cylinder cytoplazmatyczny. Na wierzchu błony zewnętrznej znajduje się przezroczysta osłona o charakterze glikozaminoglikanu. Pod zewnętrzną błoną ściany komórkowej znajdują się włókienka, które owijają się wokół cylindra cytoplazmatycznego, nadając bakteriom spiralny kształt. Włókna są przyczepione do końców komórki i skierowane ku sobie. Liczba i rozmieszczenie włókienek różni się w zależności od gatunku. Włókna biorą udział w ruchu krętków, nadając komórkom ruch obrotowy, zginający i translacyjny. W tym przypadku krętki tworzą pętle, loki i zagięcia, które nazywane są lokami wtórnymi. Krętki słabo dostrzegają barwniki. Malowane są najczęściej według Romanowskiego-Giemsy lub posrebrzane. Na żywo
Postać krętka bada się za pomocą mikroskopii z kontrastem fazowym lub mikroskopii ciemnego pola.
Krętki reprezentowane są przez trzy rodzaje patogenne dla człowieka: Treponema, Borrelia, Leptospira.
Treponema(rodzaj Treponema) mają wygląd cienkich, skręconych korkociągiem nitek z 8-12 jednolitymi małymi lokami. Wokół protoplastu krętka znajdują się 3-4 włókienka (wici). Cytoplazma zawiera włókna cytoplazmatyczne. Przedstawiciele patogenni są T. pallidum- czynnik sprawczy kiły, T. pertenue- czynnik sprawczy choroby tropikalnej ziewa. Występują także saprofity - mieszkańcy ludzkiej jamy ustnej i muł zbiorników wodnych.
Borrelia(rodzaj Borrelia), w przeciwieństwie do krętków są dłuższe, mają 3-8 dużych loków i 7-20 włókienek. Należą do nich czynnik wywołujący nawracającą gorączkę (B. recurrentis) i patogeny boreliozy (V. burgdorferi) i inne choroby.
Leptospira(rodzaj Leptospira) Mają płytkie i częste loki w postaci skręconej liny. Końce tych krętków są zakrzywione jak haczyki ze zgrubieniami na końcach. Tworząc wtórne loki, przyjmują kształt liter S lub C; mają dwa włókienka osiowe. Przedstawiciel patogenny L. interrogans powoduje leptospirozę po spożyciu z wodą lub jedzeniem, co prowadzi do krwotoku i żółtaczki.
Riketsje mają metabolizm niezależny od komórki gospodarza, jednak możliwe jest, że w celu rozmnażania otrzymują od komórki gospodarza związki wysokoenergetyczne. W rozmazach i tkankach są one barwione według Romanovsky-Giemsa, według Macchiavello-Zdrodovsky (riketsje są czerwone, a zakażone komórki są niebieskie).
U ludzi riketsje powodują epidemię tyfusu plamistego. (R. prowazekii), Riketsjoza przenoszona przez kleszcze (R. sibirica), gorączka plamista Gór Skalistych (R. rickettsii) i inne riketsjozy.
Struktura ich ściany komórkowej przypomina strukturę bakterii Gram-ujemnych, chociaż istnieją różnice. Nie zawiera typowego peptydoglikanu: całkowicie brakuje mu kwasu N-acetylomuramowego. Ściana komórkowa składa się z podwójnej błony zewnętrznej, która zawiera lipopolisacharydy i białka. Pomimo braku peptydoglikanu ściana komórkowa chlamydii jest sztywna. Cytoplazma komórki jest ograniczona przez wewnętrzną błonę cytoplazmatyczną.
Główną metodą wykrywania chlamydii jest barwienie Romanovsky-Giemsa. Kolor zależy od etapu cyklu życiowego: ciała elementarne wydają się fioletowe na tle niebieskiej cytoplazmy komórki, ciała siatkowe wydają się niebieskie.
U ludzi chlamydia powoduje uszkodzenie oczu (jaglica, zapalenie spojówek), dróg moczowo-płciowych, płuc itp.
Promieniowce- bakterie Gram-dodatnie rozgałęzione, nitkowate lub w kształcie pałeczki. Jego nazwa (z greckiego. akt- Ray, mykes- grzyb) otrzymali w wyniku tworzenia się w dotkniętych tkankach druz - granulek ściśle splecionych nici w postaci
promienie rozciągające się od środka i kończące się kolbowatymi zgrubieniami. Promieniowce, podobnie jak grzyby, tworzą grzybnię - nitkowate, przeplatające się komórki (strzępki). Tworzą grzybnię substratową, która powstaje w wyniku wrastania komórek do pożywki oraz grzybnię powietrzną, która rośnie na powierzchni pożywki. Promieniowce mogą dzielić się poprzez fragmentację grzybni na komórki podobne do bakterii pałeczek i bakterii kokosowych. Na strzępkach powietrznych promieniowców powstają zarodniki, które służą do rozmnażania. Zarodniki promieniowca zwykle nie są odporne na ciepło.
Wspólną gałąź filogenetyczną z promieniowcami tworzą tzw. promieniowce nocardiform (nocardioform) - zbiorowa grupa bakterii w kształcie pręcików o nieregularnym kształcie. Ich poszczególni przedstawiciele tworzą rozgałęzione formy. Należą do nich bakterie z rodzaju Corynebacterium, Mycobacterium, Nocardia itp. Promieniowce Nocardi-podobne wyróżniają się obecnością w ścianie komórkowej cukrów arabinozy, galaktozy, a także kwasów mykolowych i dużych ilości kwasów tłuszczowych. Kwasy mykolowe i lipidy ścian komórkowych decydują o kwasoodporności bakterii, zwłaszcza prątków gruźlicy i trądu (wybarwione według Ziehl-Neelsena są czerwone, a niekwasoodporne bakterie i elementy tkanek, plwocina mają kolor niebieski).
Patogenne promieniowce powodują promienicę, nocardia - nokardiozę, prątki - gruźlicę i trąd, maczugowców - błonicę. Saprofityczne formy promieniowców i nocardiform promieniowców są szeroko rozpowszechnione w glebie, wiele z nich jest producentami antybiotyków.
Mykoplazmy- małe bakterie (0,15-1 µm), otoczone jedynie błoną cytoplazmatyczną zawierającą sterole. Należą do klasy Mollicutes. Ze względu na brak ściany komórkowej mykoplazmy są wrażliwe osmotycznie. Mają różne kształty: kokosowy, nitkowaty, w kształcie kolby. Formy te są widoczne pod mikroskopem z kontrastem fazowym czystych kultur mykoplazm. Na gęstej pożywce mykoplazmy tworzą kolonie przypominające jajka sadzone: środkowa nieprzezroczysta część zanurzona w pożywce i półprzezroczysty obwód w kształcie koła.
Mykoplazmy powodują atypowe zapalenie płuc u ludzi (Mycoplasma pneumoniae) i uszkodzenia dróg moczowo-płciowych
(M. hominis itd.). Mykoplazmy powodują choroby nie tylko u zwierząt, ale także u roślin. Przedstawiciele niepatogenni są również dość rozpowszechnieni.
2.3. Struktura i klasyfikacja grzybów
Grzyby należą do tej domeny Eukarya, Królestwo Grzyby (Mycota, Mycetes). Ostatnio grzyby i pierwotniaki zostały podzielone na odrębne królestwa: królestwo Eumycota(prawdziwe grzyby), królestwo Chromista i królestwo Pierwotniaki. Niektóre mikroorganizmy wcześniej uważane za grzyby lub pierwotniaki zostały przeniesione do nowego królestwa Chromista(płyty chromowane). Grzyby to wielokomórkowe lub jednokomórkowe, niefotosyntetyczne (wolne od chlorofilu) mikroorganizmy eukariotyczne o grubej ścianie komórkowej. Mają jądro z otoczką jądrową, cytoplazmę z organellami, błonę cytoplazmatyczną i wielowarstwową sztywną ścianę komórkową składającą się z kilku rodzajów polisacharydów (mannany, glukany, celuloza, chityna), a także białka, lipidów itp. Niektóre grzyby tworzą kapsułka. Błona cytoplazmatyczna zawiera glikoproteiny, fosfolipidy i ergosterole (w przeciwieństwie do cholesterolu, głównego sterolu tkanek ssaków). Większość grzybów to tlenowce obligatoryjne lub fakultatywne.
Grzyby są szeroko rozpowszechnione w przyrodzie, zwłaszcza w glebie. Niektóre grzyby przyczyniają się do produkcji chleba, serów, produktów na bazie kwasu mlekowego i alkoholu. Inne grzyby wytwarzają antybiotyki przeciwdrobnoustrojowe (np. penicylinę) i leki immunosupresyjne (np. cyklosporynę). Grzyby są wykorzystywane przez genetyków i biologów molekularnych do modelowania różnych procesów. Grzyby fitopatogenne powodują znaczne szkody w rolnictwie, powodując choroby grzybowe roślin zbożowych i zbóż. Infekcje wywołane przez grzyby nazywane są grzybicami. Istnieją grzyby strzępkowe i drożdżowe.
Grzyby strzępkowe (pleśniowe) lub Hyphomycetes składają się z cienkich nitek o grubości 2-50 mikronów, zwanych strzępkami, które są wplecione w grzybnię lub grzybnię (pleśń). Ciało grzyba nazywa się plechą. Istnieją hyphomycetes demacium (pigmentowane - brązowe lub czarne) i szkliste (niepigmentowane). Strzępki wrastające w podłoże odżywcze odpowiadają za odżywianie grzyba i nazywane są strzępkami wegetatywnymi. Hyphae, ra-
stojące nad powierzchnią podłoża nazywane są strzępkami powietrznymi lub rozrodczymi (odpowiedzialnymi za rozmnażanie). Kolonie mają puszysty wygląd dzięki grzybni powietrznej.
Wyróżnia się grzyby niższe i wyższe: strzępki grzybów wyższych oddzielone są przegrodami, czyli przegrodami z otworami. Strzępki grzybów niższych nie mają przegród, są to komórki wielojądrzaste zwane koenocytami (z gr. koenos- pojedynczy, pospolity).
Grzyby drożdżowe (drożdże) są reprezentowane głównie przez pojedyncze owalne komórki o średnicy 3-15 mikronów, a ich kolonie, w przeciwieństwie do grzybów strzępkowych, mają zwarty wygląd. W zależności od rodzaju rozmnażania płciowego są one rozprowadzane wśród wyższych grzybów - ascomycetes i basidiomycetes. Podczas rozmnażania bezpłciowego drożdże pączkują lub dzielą się. Mogą tworzyć pseudostrzępki i fałszywą grzybnię (pseudomycelium) w postaci łańcuchów wydłużonych komórek - „kiełbasek”. Grzyby podobne do drożdży, ale nie mające płciowej metody rozmnażania, nazywane są drożdżopodobnymi. Rozmnażają się wyłącznie bezpłciowo - przez pączkowanie lub rozszczepienie. Pojęcia „grzyby drożdżopodobne” często utożsamiane są z pojęciem „drożdże”.
Wiele grzybów charakteryzuje się dymorfizmem - zdolnością do wzrostu strzępków (grzybni) lub drożdżopodobnych, w zależności od warunków uprawy. W zakażonym organizmie rosną w postaci komórek drożdżopodobnych (faza drożdżowa), a na pożywce tworzą strzępki i grzybnię. Dymorfizm jest powiązany z czynnikiem temperaturowym: w temperaturze pokojowej tworzy się grzybnia, a w temperaturze 37 ° C (w temperaturze ciała ludzkiego) tworzą się komórki drożdżopodobne.
Grzyby rozmnażają się płciowo lub bezpłciowo. Rozmnażanie płciowe grzybów następuje wraz z tworzeniem się gamet, zarodników płciowych i innych form płciowych. Formy płciowe nazywane są teleomorfami.
Rozmnażanie bezpłciowe grzybów następuje wraz z utworzeniem odpowiednich form zwanych anamorfami. Takie rozmnażanie następuje poprzez pączkowanie, fragmentację strzępek i zarodników bezpłciowych. Endogenne zarodniki (sporangiospory) dojrzewają wewnątrz okrągłej struktury - zarodni. Na końcach strzępek owocujących, tzw. konidioforach, tworzą się egzogenne zarodniki (konidia).
Istnieje wiele konidiów. Arthroconidia (arthrospory), czyli talokonidia, powstają w wyniku jednolitej przegrody i rozczłonkowania strzępek, a blastoconidia powstają w wyniku pączkowania. Małe konidia jednokomórkowe nazywane są mikrokonidiami, duże konidia wielokomórkowe nazywane są makrokonidiami. Bezpłciowe formy grzybów obejmują również chlamydokonidie lub chlamydospory (grubościenne duże komórki spoczynkowe lub kompleks małych komórek).
Są grzyby doskonałe i niedoskonałe. Grzyby doskonałe mają metodę rozmnażania płciowego; obejmują one zygomycetes (Zygomykota), workowce (Ascomycota) i podstawczaki (Podstawczaki). Grzyby niedoskonałe rozmnażają się wyłącznie bezpłciowo; Należą do nich formalny konwencjonalny typ/grupa grzybów - Deuteromycetes (Deiteromykota).
Zygomycetes należą do grzybów niższych (grzybnia nieseptyczna). Należą do nich przedstawiciele rodzajów Mucor, Rhizopus, Rhizomucor, Absidia, Basidiobolus, Conidiobolus. Ukazuje się w glebie i powietrzu. Mogą powodować zygomykozę (mukoromykozę) płuc, mózgu i innych narządów ludzkich.
Podczas bezpłciowego rozmnażania zygomycetes na strzępkach owocujących (sporangioforach) tworzy się zarodnia - kuliste zgrubienie ze skorupą zawierającą liczne sporangiospory (ryc. 2.6, 2.7). Rozmnażanie płciowe u zygomycetes odbywa się za pomocą zygospor.
Ascomycetes (grzyby torbacze) mają grzybnię z przegrodą (z wyjątkiem drożdży jednokomórkowych). Swoją nazwę wzięli od głównego narządu owocującego - kaletki lub worka, zawierającego 4 lub 8 haploidalnych zarodników płciowych (askospor).
Ascomycetes obejmują indywidualnych przedstawicieli (teleomorfy) rodzajów Aspergillus I Penicillium. Większość rodzajów grzybów Aspergillus, Penicillium są anamorficzne, tj. rozmnażają się tylko bezradnie
 Ryż. 2.6. Grzyby z rodzaju Mucor(rysunek: A.S. Bykov)
Ryż. 2.6. Grzyby z rodzaju Mucor(rysunek: A.S. Bykov)
 Ryż. 2.7. Grzyby z rodzaju Rhizopus. Rozwój zarodni, sporangiospor i ryzoidów
Ryż. 2.7. Grzyby z rodzaju Rhizopus. Rozwój zarodni, sporangiospor i ryzoidów
bezpośrednio przy pomocy zarodników bezpłciowych - konidiów (ryc. 2.8, 2.9) i ze względu na tę cechę należy je klasyfikować jako grzyby niedoskonałe. W grzybach z rodzaju Aspergillus na końcach strzępek owocujących, konidioforów, znajdują się zgrubienia - sterigmata, fialidy, na których tworzą się łańcuchy konidiów („pleśń wodna”).
W grzybach z rodzaju Penicillium(frędzel) strzępka owocująca przypomina pędzel, ponieważ tworzą się z niej zgrubienia (na konidioforze), rozgałęziające się na mniejsze struktury - sterigmata, fialidy, na których znajdują się łańcuchy konidiów. Niektóre gatunki Aspergillus mogą powodować aspergilozę i aflatoksykozę, a Penicillium może powodować penicyliozę.
Przedstawiciele workowców są teleomorfami tego rodzaju Trichophyton, Microsporum, Histoplasma, Blastomyces, jak i dreszcze
 Ryż. 2.8. Grzyby z rodzaju Penicillium. Z fialid wystają łańcuchy konidiów
Ryż. 2.8. Grzyby z rodzaju Penicillium. Z fialid wystają łańcuchy konidiów
 Ryż. 2.9. Grzyby z rodzaju Aspergillus fumigatus. Z fialid wystają łańcuchy konidiów
Ryż. 2.9. Grzyby z rodzaju Aspergillus fumigatus. Z fialid wystają łańcuchy konidiów
Basidiomycetes obejmują grzyby kapeluszowe. Mają przegrodową grzybnię i tworzą zarodniki płciowe - bazydiospory, oddzielając się od podstawki - końcowej komórki grzybni, homologicznej do worka. Basidiomycetes obejmują niektóre drożdże, takie jak teleomorfy Cryptococcus neoformans.
Deuteromycetes to grzyby niedoskonałe (Grzyby niedoskonałe, grzyby anamorficzne, grzyby konidialne). Jest to warunkowy, formalny takson grzybów, łączący grzyby, które nie rozmnażają się płciowo. Ostatnio zamiast terminu „deuteromycetes” zaproponowano określenie „grzyby mitosporne” - grzyby rozmnażające się przez zarodniki niepłciowe, tj. przez mitozę. Po ustaleniu faktu rozmnażania płciowego niedoskonałych grzybów, są one przenoszone do jednego ze znanych typów - Ascomycota Lub Podstawczaki nadanie nazwie formy teleomorficznej. Deuteromycetes mają grzybnię przegrodową i rozmnażają się wyłącznie poprzez bezpłciowe tworzenie konidiów. Deuteromycetes obejmują niedoskonałe drożdże (grzyby drożdżopodobne), na przykład niektóre grzyby z rodzaju Candida, wpływające na skórę, błony śluzowe i narządy wewnętrzne (kandydoza). Mają owalny kształt, średnicę 2-5 mikronów, dzielą się przez pączkowanie, tworzą pseudostrzępki (pseudomycelium) w postaci łańcuchów wydłużonych komórek, a czasem tworzą strzępki. Dla Candida albicans charakterystyczne jest tworzenie chlamydospor (ryc. 2.10). Do Deuteromycetes zaliczają się także inne grzyby, które nie mają płciowego sposobu rozmnażania, należące do tego rodzaju Epidermophyton, Coccidioides, Paracoccidioides, Sporothrix, Aspergillus, Phialophora, Fonsecaea, Exophiala, Cladophialophora, Bipolaris, Exerohilum, Wangiella, Alrernaria itd.
 Ryż. 2.10. Grzyby z rodzaju Candida albicans(rysunek: A.S. Bykov)
Ryż. 2.10. Grzyby z rodzaju Candida albicans(rysunek: A.S. Bykov)
2.4. Budowa i klasyfikacja pierwotniaków
Najprostsze należą do domeny Eukarya, Królestwo zwierząt (Zwierzęta), podkrólestwo Pierwotniaki. Niedawno zaproponowano przypisanie pierwotniaków do rangi królestwa Pierwotniaki.
Komórka pierwotniaka jest otoczona błoną (błonką) - analogiem błony cytoplazmatycznej komórek zwierzęcych. Ma jądro z otoczką jądrową i jąderkiem, cytoplazmę zawierającą retikulum endoplazmatyczne, mitochondria, lizosomy i rybosomy. Rozmiary pierwotniaków wahają się od 2 do 100 mikronów. Po zabarwieniu według Romanovsky-Giemsa jądro pierwotniaka jest czerwone, a cytoplazma niebieska. Pierwotniaki poruszają się za pomocą wici, rzęsek lub pseudopodiów, niektóre z nich mają wakuole trawienne i kurczliwe (wydalnicze). Mogą żerować w wyniku fagocytozy lub tworzenia specjalnych struktur. W zależności od rodzaju odżywiania dzieli się je na heterotrofy i autotrofy. Wiele pierwotniaków (ameba czerwonkowa, Giardia, Trichomonas, Leishmania, Balantidia) może rosnąć na pożywkach zawierających natywne białka i aminokwasy. Do ich hodowli wykorzystywane są także kultury komórkowe, zarodki kurze i zwierzęta laboratoryjne.
Pierwotniaki rozmnażają się bezpłciowo – poprzez podział podwójny lub wielokrotny (schizogonia), a niektóre także płciowo (sporogonia). Niektóre pierwotniaki rozmnażają się pozakomórkowo (Giardia), inne zaś wewnątrzkomórkowo (Plasmodium, Toxoplasma, Leishmania). Cykl życiowy pierwotniaków charakteryzuje się etapami - tworzeniem się stadium trofozoitu i stadium cysty. Cysty to stadia uśpione, odporne na zmiany temperatury i wilgotności. Cysty są odporne na kwasy Sarcocystis, Cryptosporidium I Isospora.
Wcześniej pierwotniaki wywołujące choroby u ludzi reprezentowane były przez 4 typy 1 ( Sarcomastigophora, Apicomplexa, Ciliophora, Microspora). Te typy zostały ostatnio przeklasyfikowane na większą liczbę i wyłoniły się nowe królestwa - Pierwotniaki I Chromista(Tabela 2.2). Do nowego królestwa Chromista(chromovics) obejmowały niektóre pierwotniaki i grzyby (blastocysty, oomycetes i Rhinosporidium seeberi). Królestwo Pierwotniaki obejmuje ameby, wiciowce, sporozoany i orzęski. Dzielą się na różne typy, wśród których są ameby, wiciowce, sporozoany i orzęski.
Tabela 2.2. Przedstawiciele królestw Pierwotniaki I Chromista, o znaczeniu medycznym
 1 Typ Sarkomastygofora składał się z podtypów Sarkodyna I Mastigofora. Podtyp Sarkodyna(sarcodaceae) obejmowały amebę czerwonkową i jej podtyp Mastigofora(wiciowce) - trypanosomy, leishmania, lamblia i trichomonas. Typ Apikompleks klasa uwzględniona Sporozoa(zarodniki), do których zaliczają się plazmodia malarii, toksoplazma, cryptosporidium itp. Typ Ciliofora obejmuje balantidię i typ Mikrospora- mikrosporydia.
1 Typ Sarkomastygofora składał się z podtypów Sarkodyna I Mastigofora. Podtyp Sarkodyna(sarcodaceae) obejmowały amebę czerwonkową i jej podtyp Mastigofora(wiciowce) - trypanosomy, leishmania, lamblia i trichomonas. Typ Apikompleks klasa uwzględniona Sporozoa(zarodniki), do których zaliczają się plazmodia malarii, toksoplazma, cryptosporidium itp. Typ Ciliofora obejmuje balantidię i typ Mikrospora- mikrosporydia.
Koniec stołu. 2.2
 Ameby obejmują czynnik wywołujący ludzką amebozę - czerwonkę amebową (Entamoeba histolytica), wolno żyjące i niepatogenne ameby (ameba jelitowa itp.). Ameby rozmnażają się binarnie, bezpłciowo. Ich cykl życiowy składa się z etapu trofozoitu (rosnącej, ruchliwej komórki, słabo stabilnej) i stadium cysty. Trofozoity poruszają się za pomocą pseudopodiów, które wychwytują i zanurzają składniki odżywcze w cytoplazmie. Z
Ameby obejmują czynnik wywołujący ludzką amebozę - czerwonkę amebową (Entamoeba histolytica), wolno żyjące i niepatogenne ameby (ameba jelitowa itp.). Ameby rozmnażają się binarnie, bezpłciowo. Ich cykl życiowy składa się z etapu trofozoitu (rosnącej, ruchliwej komórki, słabo stabilnej) i stadium cysty. Trofozoity poruszają się za pomocą pseudopodiów, które wychwytują i zanurzają składniki odżywcze w cytoplazmie. Z
Trofozoit tworzy cystę odporną na działanie czynników zewnętrznych. Po dostaniu się do jelita zamienia się w trofozoit.
Wiciowce charakteryzują się obecnością wici: Leishmania ma jedną wici, Trichomonas ma 4 wolne wici i jedną wici połączone z krótką falującą błoną. Oni są:
Wiciowce krwi i tkanek (Leishmania - czynniki wywołujące leiszmaniozę; Trypanosomy - czynniki powodujące śpiączkę i chorobę Chagasa);
Wiciowce jelitowe (giardia - czynnik wywołujący lambliozę);
Wiciowce dróg moczowo-płciowych (Trichomonas pochwylis – czynnik wywołujący rzęsistkowicę).
Rzęskowe są reprezentowane przez balantidia, które atakują ludzką okrężnicę (czerwonka balantidioza). Balantidia mają stadium trofozoitu i cysty. Trofozoit jest mobilny, ma liczne rzęski, cieńsze i krótsze niż wici.
2.5. Struktura i klasyfikacja wirusów
Wirusy to najmniejsze drobnoustroje należące do królestwa Wirusy(od łac. wirus- I). Nie mają struktury komórkowej i składają się
Strukturę wirusów, ze względu na ich niewielkie rozmiary, bada się za pomocą mikroskopii elektronowej obu wirionów i ich ultracienkich przekrojów. Wielkość wirusów (wirionów) określa się bezpośrednio za pomocą mikroskopii elektronowej lub pośrednio poprzez ultrafiltrację przez filtry o znanej średnicy porów lub ultrawirowanie. Wielkość wirusów mieści się w zakresie od 15 do 400 nm (1 nm równa się 1/1000 μm): do małych wirusów, których wielkość jest zbliżona do wielkości rybosomów, zaliczają się parwowirusy i wirus polio, a największe to wirus ospy wietrznej (350 nm). Wirusy różnią się kształtem swoich wirionów, które mają postać pręcików (wirus mozaiki tytoniowej), kul (wirus wścieklizny), kulek (wirusy poliomyelitis, HIV), nici (filowirusy), plemników (wiele bakteriofagów).
Wirusy zadziwiają wyobraźnią różnorodnością budowy i właściwości. W przeciwieństwie do genomów komórkowych, które zawierają jednolity dwuniciowy DNA, genomy wirusowe są niezwykle zróżnicowane. Istnieją wirusy DNA i RNA, które są haploidalne, tj. mają jeden zestaw genów. Tylko retrowirusy mają diploidalny genom. Genom wirusów zawiera od 6 do 200 genów i jest reprezentowany przez różne typy kwasów nukleinowych: dwuniciowe, jednoniciowe, liniowe, koliste, fragmentowane.
Wśród wirusów jednoniciowych RNA rozróżnia się genomowy RNA o nici dodatniej i RNA o nici ujemnej (polaryzacja RNA). Nić dodatnia (nić dodatnia) RNA tych wirusów, oprócz funkcji genomowej (dziedzicznej), pełni funkcję informacyjnego lub informacyjnego RNA (mRNA lub mRNA); jest matrycą do syntezy białek na rybosomach zakażonej komórki. Plusniciowy RNA jest zakaźny: wprowadzony do wrażliwych komórek może wywołać proces zakaźny.
ces. Nić ujemna (nić ujemna) wirusów RNA pełni jedynie funkcję dziedziczną; Do syntezy białek na nici ujemnej RNA syntetyzowana jest nić komplementarna. Niektóre wirusy mają ambipolarny genom RNA (ambisens z języka greckiego Ambi- po obu stronach podwójna komplementarność), tj. zawiera plus i minus segmenty RNA.
Istnieją proste wirusy (na przykład wirus zapalenia wątroby typu A) i złożone wirusy (na przykład wirusy grypy, opryszczki, koronawirusy).
Wirusy proste lub bezotoczkowe mają jedynie kwas nukleinowy związany ze strukturą białkową zwaną kapsydem (od łac. capsa- sprawa). Białka związane z kwasem nukleinowym nazywane są nukleoproteinami, a połączenie wirusowych białek kapsydu z wirusowym kwasem nukleinowym nazywa się nukleokapsydem. Niektóre proste wirusy mogą tworzyć kryształy (np. wirus pryszczycy).
Kapsyd zawiera powtarzające się podjednostki morfologiczne - kapsomery, złożone z kilku polipeptydów. Kwas nukleinowy wirionu wiąże się z kapsydem, tworząc nukleokapsyd. Kapsyd chroni kwas nukleinowy przed degradacją. W prostych wirusach kapsyd bierze udział w przyłączaniu (adsorpcji) do komórki gospodarza. Proste wirusy opuszczają komórkę w wyniku jej zniszczenia (lizy).
Wirusy złożone, czyli otoczkowe (ryc. 2.11), oprócz kapsydu, mają podwójną otoczkę lipoproteinową (synonim: superkapsyd lub peplos), która jest nabywana przez pączkowanie wirionu przez błonę komórkową, na przykład przez błona plazmatyczna, błona jądrowa lub błona siateczki śródplazmatycznej. Otoczka wirusa zawiera kolce glikoproteinowe,
lub kolce, peplomery. Zniszczenie otoczki eterem i innymi rozpuszczalnikami inaktywuje złożone wirusy. Pod otoczką niektórych wirusów znajduje się białko macierzy (białko M).
Wiriony mają spiralną, dwudziestościenną (sześcienną) lub złożoną symetrię kapsydu (nukleokapsyd). Symetria helikalna wynika ze spiralnej struktury nukleokapsydu (na przykład w wirusach grypy, koronawirusach): kapsomery są ułożone spiralnie wraz z kwasem nukleinowym. Symetria ikozaedryczna wynika z utworzenia izometrycznie pustego ciała z kapsydu zawierającego wirusowy kwas nukleinowy (na przykład w wirusie opryszczki).
Kapsyd i otoczka (superkapsyd) chronią wiriony przed wpływami środowiska i determinują selektywne oddziaływanie (adsorpcję) ich białek receptorowych z określonymi
 Ryż. 2.11. Struktura wirusów otoczkowych z kapsydem dwudziestościennym (a) i helikalnym (b).
Ryż. 2.11. Struktura wirusów otoczkowych z kapsydem dwudziestościennym (a) i helikalnym (b).
komórek, a także właściwości antygenowe i immunogenne wirionów.
Wewnętrzne struktury wirusów nazywane są rdzeniem. W adenowirusach rdzeń składa się z białek histonopodobnych związanych z DNA, w reowirusach - z białek wewnętrznego kapsydu.
Laureat Nagrody Nobla D. Baltimore zaproponował system klasyfikacji Baltimore oparty na mechanizmie syntezy mRNA. Klasyfikacja ta dzieli wirusy na 7 grup (Tabela 2.3). Międzynarodowy Komitet Taksonomii Wirusów (telewizja telewizyjna) przyjęła uniwersalny system klasyfikacji, który wykorzystuje kategorie taksonomiczne, takie jak rodzina (imię kończące się na wirusy), podrodzina (nazwa kończy się na virinae), rodzaj (nazwa kończy się na wirus). Gatunek wirusa nie otrzymał nazwy dwumianowej, podobnie jak bakterie. Wirusy są klasyfikowane według rodzaju kwasu nukleinowego (DNA lub RNA), jego struktury i liczby nici. Mają dwuniciowe lub jednoniciowe kwasy nukleinowe; dodatnia (+), ujemna (-) polaryzacja kwasu nukleinowego lub mieszana polaryzacja kwasu nukleinowego, dwubiegunowa (+, -); liniowy lub kolisty kwas nukleinowy; fragmentowany lub niefragmentowany kwas nukleinowy. Brane są również pod uwagę wielkość i morfologia wirionów, liczba kapsomerów i rodzaj symetrii nukleokapsydu, obecność otoczki (superkapsydu), wrażliwość na eter i dezoksycholan, miejsce rozmnażania w komórce, właściwości antygenowe itp. na konto.
Tabela 2.3. Główne wirusy o znaczeniu medycznym
 Kontynuacja tabeli. 2.3
Kontynuacja tabeli. 2.3
 Koniec stołu. 2.3
Koniec stołu. 2.3
 Wirusy infekują zwierzęta, bakterie, grzyby i rośliny. Będąc głównymi czynnikami sprawczymi chorób zakaźnych człowieka, wirusy biorą również udział w procesach karcynogenezy i mogą być przenoszone na różne sposoby, w tym przez łożysko (wirus różyczki, wirus cytomegalii itp.), Wpływając na ludzki płód. Mogą również prowadzić do powikłań poinfekcyjnych - rozwoju zapalenia mięśnia sercowego, zapalenia trzustki, niedoboru odporności itp.
Wirusy infekują zwierzęta, bakterie, grzyby i rośliny. Będąc głównymi czynnikami sprawczymi chorób zakaźnych człowieka, wirusy biorą również udział w procesach karcynogenezy i mogą być przenoszone na różne sposoby, w tym przez łożysko (wirus różyczki, wirus cytomegalii itp.), Wpływając na ludzki płód. Mogą również prowadzić do powikłań poinfekcyjnych - rozwoju zapalenia mięśnia sercowego, zapalenia trzustki, niedoboru odporności itp.
Oprócz wirusów do niekomórkowych form życia zaliczają się priony i wiroidy. Wiroidy to małe cząsteczki kolistego, superskręconego RNA, które nie zawierają białka i powodują choroby roślin. Patologiczne priony to zakaźne cząstki białkowe, które powodują szczególne choroby konformacyjne w wyniku zmian w strukturze normalnego komórkowego białka prionowego ( PrP c), który występuje w organizmie zwierząt i ludzi. PrP z pełni funkcje regulacyjne. Jest kodowany przez normalny gen prionu (gen PrP), zlokalizowany na krótkim ramieniu ludzkiego chromosomu 20. Choroby prionowe występują w postaci pasażowalnych encefalopatii gąbczastych (choroba Creutzfeldta-Jakoba, kuru itp.). W tym przypadku białko prionowe przybiera inną, zakaźną formę, oznaczoną jako PrP sc(sc z trzęsawka- trzęsawka to infekcja prionowa owiec i kóz). To zakaźne białko prionowe ma wygląd włókienek i różni się od normalnego białka prionowego swoją trzeciorzędową lub czwartorzędową strukturą.
Zadania do samodzielnego przygotowania (samokontroli)
A. Wskaż mikroorganizmy należące do prokariotów:
2. Wirusy.
3. Bakterie.
4. Priony.
B. Zwróć uwagę na charakterystyczne cechy komórki prokariotycznej:
1. Rybosomy 70S.
2. Obecność peptydoglikanu w ścianie komórkowej.
3. Obecność mitochondriów.
4. Diploidalny zestaw genów.
W. Oznacz składniki peptydoglikanu:
1. Kwasy tejchojowe.
2. N-acetyloglukozamina.
3. Lipopolisaryd.
4. Tetrapeptyd.
G. Zwróć uwagę na cechy strukturalne ściany komórkowej bakterii Gram-ujemnych:
1. Kwas mezodiaminopimelinowy.
2. Kwasy tejchojowe.
4. Białka Porin.
D. Wymień funkcje zarodników u bakterii:
1. Zachowanie gatunku.
2. Odporność na ciepło.
3. Rozprowadzenie podłoża.
4. Powielanie.
1. Riketsja.
2. Promieniowce.
3. Krętki.
4. Chlamydie.
I. Wymień cechy promieniowców:
1. Mają termolabilne zarodniki.
2. Bakterie Gram-dodatnie.
3. Nie ma ściany komórkowej.
4. Mają zawiły kształt.
Z. Wymień cechy krętków:
1. Bakterie Gram-ujemne.
2. Mają aparat włóknisty silnikowy.
3. Mają zawiły kształt.
I. Wymień pierwotniaki posiadające kompleks wierzchołkowy umożliwiający im wniknięcie do komórki:
1. Plazmodium malarii.
3. Toksoplazma.
4. Kryptosporydium.
DO. Wymień charakterystyczną cechę złożonych wirusów:
1. Dwa rodzaje kwasów nukleinowych.
2. Obecność błony lipidowej.
3. Podwójny kapsyd.
4. Obecność białek niestrukturalnych. L. Zaznacz wyższe grzyby:
1. Mucor.
2. Candida.
3. Penicillium.
4. Aspergillus.
Mikroorganizmy (mikroorganizmy) to organizmy jednokomórkowe o wielkości mniejszej niż 0,1 mm, których nie można zobaczyć gołym okiem. Należą do nich bakterie, mikroalgi, niektóre niższe grzyby nitkowate, drożdże i pierwotniaki (ryc. 1). Bada je mikrobiologia.
Ryż. 1. Obiekty mikrobiologiczne.
Na ryc. 2. Można zobaczyć niektórych przedstawicieli pierwotniaków jednokomórkowych. Czasami przedmiotem tej nauki są najbardziej prymitywne organizmy na Ziemi - wirusy, które nie mają struktury komórkowej i są kompleksami kwasów nukleinowych (materiału genetycznego) i białka. Częściej są one izolowane w zupełnie odrębnym kierunku studiów (wirusologia), ponieważ mikrobiologia jest raczej nastawiona na badanie mikroskopijnych organizmów jednokomórkowych.

Ryż. 2. Poszczególni przedstawiciele jednokomórkowych eukariontów (pierwotniaków).
Nauki algologiczne i mykologiczne, które badają odpowiednio glony i grzyby, to odrębne dyscypliny, które pokrywają się z mikrobiologią w badaniu mikroskopijnych obiektów żywych. Bakteriologia to prawdziwa dziedzina mikrobiologii. Nauka ta zajmuje się wyłącznie badaniem mikroorganizmów prokariotycznych (ryc. 3).

Ryż. 3. Schemat komórki prokariotycznej.
W przeciwieństwie do eukariontów, do których zaliczają się wszystkie organizmy wielokomórkowe, a także pierwotniaki, mikroskopijne algi i grzyby, prokarioty nie mają utworzonego jądra zawierającego materiał genetyczny i prawdziwych organelli (trwałych wyspecjalizowanych struktur komórki).
Do prokariotów zaliczają się bakterie prawdziwe i archeony, które według współczesnej klasyfikacji określane są jako domeny (superkrólestwa) Archaea i Eubacteria (ryc. 4).

Ryż. 4. Dziedziny współczesnej klasyfikacji biologicznej.
Cechy strukturalne bakterii
Bakterie są ważnym ogniwem obiegu substancji w przyrodzie, rozkładają pozostałości roślinne i zwierzęce, oczyszczają zbiorniki wodne zanieczyszczone materią organiczną, modyfikują związki nieorganiczne. Bez nich życie na ziemi nie mogłoby istnieć. Mikroorganizmy te są rozmieszczone wszędzie, w glebie, wodzie, powietrzu, organizmach zwierzęcych i roślinnych.
Bakterie różnią się następującymi cechami morfologicznymi:
- Kształt komórki (okrągły, w kształcie pręta, nitkowaty, zawiły, spiralny, a także różne opcje przejściowe i konfiguracja w kształcie gwiazdy).
- Obecność urządzeń do poruszania się (nieruchomych, biczowanych, z powodu wydzielania śluzu).
- Artykulacja komórek między sobą (izolowane, połączone w postaci par, granulek, form rozgałęzionych).
Wśród struktur utworzonych przez bakterie okrągłe (cocci) znajdują się komórki, które po podziale łączą się w pary, a następnie rozpadają się na pojedyncze formacje (mikrokoki) lub pozostają cały czas razem (diplokoki). Kwadratowa struktura czterech komórek jest utworzona przez tetrakoki, łańcuch przez paciorkowce, ziarnistość 8-64 jednostek przez sarcina i skupiska przez gronkowce.
Bakterie pałeczkowate reprezentowane są przez różnorodne kształty ze względu na dużą zmienność długości (0,1-15 µm) i grubości (0,1-2 µm) komórki. Kształt tych ostatnich zależy także od zdolności bakterii do tworzenia zarodników – struktur o grubej otoczce, która pozwala mikroorganizmom przetrwać niesprzyjające warunki. Komórki posiadające tę zdolność nazywane są prątkami, a te pozbawione takich właściwości nazywane są po prostu bakteriami w kształcie pałeczek.
Szczególnymi modyfikacjami bakterii w kształcie pręcików są formy nitkowate (wydłużone), łańcuchy i struktury rozgałęzione. Ten ostatni jest tworzony przez promieniowce na pewnym etapie rozwoju. „Zakrzywione” pręty nazywane są bakteriami karbowanymi, spośród których izolowane są wibratory; spirilla posiadająca dwa zagięcia (15-20 µm); krętki przypominające faliste linie. Długości ich komórek wynoszą odpowiednio 1-3, 15-20 i 20-30 µm. Na ryc. Ryciny 5 i 6 przedstawiają główne formy morfologiczne bakterii, a także rodzaje ułożenia zarodników w komórce.

Ryż. 5. Podstawowe formy bakterii.

Ryż. 6. Bakterie według rodzaju umiejscowienia zarodników w komórce. 1, 4 – w środku; 2, 3, 5 – lokalizacja końcowa; 6 – z boku.
Główne struktury komórkowe bakterii: nukleoid (materiał genetyczny), rybosomy przeznaczone do syntezy białek, błona cytoplazmatyczna (część błony komórkowej), która u wielu przedstawicieli jest dodatkowo chroniona od góry, otoczka i osłonka śluzowa (ryc. 7).

Ryż. 7. Schemat komórki bakteryjnej.
Według klasyfikacji bakterii istnieje ponad 20 typów. Na przykład wyjątkowo ciepłolubne (miłośników wysokich temperatur) Aquificae, beztlenowe bakterie w kształcie pałeczki Bacteroidetes. Jednak najbardziej dominującym typem, który obejmuje różnorodnych przedstawicieli, jest Actinobacteria. Obejmuje bifidobakterie, pałeczki kwasu mlekowego i promieniowce. Wyjątkowość tego ostatniego polega na zdolności do tworzenia grzybni na pewnym etapie rozwoju.
W języku potocznym nazywa się to grzybnią. Rzeczywiście, rozgałęziające się komórki promieniowców przypominają strzępki grzybów. Pomimo tej cechy promieniowce są klasyfikowane jako bakterie, ponieważ są prokariotami. Naturalnie ich komórki są mniej podobne w budowie do grzybów.
Promieniowce (ryc. 8) są bakteriami wolno rosnącymi i dlatego nie mają zdolności konkurowania o łatwo dostępne substraty. Są zdolne do rozkładu substancji, których inne mikroorganizmy nie mogą wykorzystać jako źródła węgla, w szczególności węglowodorów ropopochodnych. Dlatego promieniowce są intensywnie badane w dziedzinie biotechnologii.
Niektórzy przedstawiciele koncentrują się na obszarach pól naftowych i tworzą specjalny filtr bakteryjny, który zapobiega przedostawaniu się węglowodorów do atmosfery. Promieniowce są aktywnymi producentami praktycznie cennych związków: witamin, kwasów tłuszczowych, antybiotyków.

Ryż. 8. Reprezentatywny promieniowiec Nocardia.
Grzyby w mikrobiologii
Przedmiotem mikrobiologii są wyłącznie grzyby pleśniowe (w szczególności rhizopus, śluz). Jak wszystkie grzyby, nie są w stanie samodzielnie syntetyzować substancji i wymagają pożywki. Grzybnia niższych przedstawicieli tego królestwa jest prymitywna, nie podzielona podziałami. Szczególną niszę w badaniach mikrobiologicznych zajmują drożdże (ryc. 9), charakteryzujące się brakiem grzybni.

Ryż. 9. Formy kolonii kultur drożdży na pożywce.
Obecnie zgromadzono wiele wiedzy na temat ich dobroczynnych właściwości. Jednakże drożdże są nadal badane pod kątem zdolności do syntezy praktycznie cennych związków organicznych i są aktywnie wykorzystywane jako organizmy modelowe w eksperymentach genetycznych. Od czasów starożytnych drożdże były wykorzystywane w procesach fermentacji. Metabolizm różni się u różnych przedstawicieli. Dlatego niektóre drożdże są bardziej odpowiednie do określonego procesu niż inne.
Na przykład Saccharomyces beticus, który jest bardziej odporny na wysokie stężenia alkoholu, wykorzystuje się do tworzenia mocnych win (do 24%). Natomiast drożdże S. cerevisiae są w stanie wytworzyć niższe stężenia etanolu. Ze względu na obszar zastosowania drożdże dzieli się na: paszowe, piekarskie, browarnicze, spirytusowe i winne.
Mikroorganizmy chorobotwórcze
Mikroorganizmy chorobotwórcze lub chorobotwórcze można znaleźć wszędzie. Oprócz dobrze znanych wirusów: grypy, zapalenia wątroby, odry, HIV itp., niebezpiecznymi mikroorganizmami są riketsje, a także paciorkowce i gronkowce, które powodują zatrucie krwi. Wśród bakterii pałeczek występuje wiele patogenów. Na przykład błonica, gruźlica, dur brzuszny (ryc. 10). Wśród pierwotniaków występuje wielu przedstawicieli mikroorganizmów niebezpiecznych dla człowieka, w szczególności plazmodium malarii, toksoplazma, leiszmania, lamblia, rzęsistki i chorobotwórcze ameby.

Ryż. 10. Zdjęcie bakterii Bacillus anthracis, która powoduje wąglika.
Wiele promieniowców nie jest niebezpiecznych dla ludzi i zwierząt. Jednak wśród prątków wywołujących gruźlicę i trąd znajduje się wielu przedstawicieli patogenów. Niektóre promieniowce inicjują chorobę taką jak promienica, której towarzyszy powstawanie ziarniniaków, a czasami wzrost temperatury ciała. Niektóre rodzaje grzybów pleśniowych są zdolne do wytwarzania substancji toksycznych dla człowieka – mikotoksyn. Na przykład niektórzy przedstawiciele rodzaju Aspergillus, Fusarium. Grzyby chorobotwórcze powodują grupę chorób zwanych grzybicami. Zatem kandydoza, czyli po prostu pleśniawka, jest wywoływana przez grzyby drożdżopodobne (ryc. 11). Są zawsze obecne w organizmie człowieka, jednak aktywują się dopiero wtedy, gdy układ odpornościowy jest osłabiony.

Ryż. 11. Grzyb Candida jest przyczyną pleśniawki.
Grzyby mogą powodować różnorodne zmiany skórne, w szczególności wszelkiego rodzaju porosty, z wyjątkiem półpaśca, który wywołuje wirus. Drożdże Malassezia, stali mieszkańcy ludzkiej skóry, mogą powodować zmniejszenie aktywności układu odpornościowego. Nie spiesz się od razu, aby umyć ręce. Drożdże i bakterie oportunistyczne w dobrym zdrowiu pełnią ważną funkcję zapobiegającą rozwojowi patogenów.
Wirusy jako przedmiot mikrobiologii
Wirusy to najbardziej prymitywne organizmy na Ziemi. W stanie wolnym nie zachodzą w nich żadne procesy metaboliczne. Dopiero po wejściu do komórki gospodarza wirusy zaczynają się namnażać. We wszystkich organizmach żywych nośnikiem materiału genetycznego jest kwas dezoksyrybonukleinowy (DNA). Tylko wśród wirusów istnieją przedstawiciele posiadający sekwencję genetyczną, taką jak kwas rybonukleinowy (RNA).
Wirusów często nie klasyfikuje się jako organizmów prawdziwie żywych.
Morfologia wirusów jest bardzo zróżnicowana (ryc. 12). Zazwyczaj ich średnice mieszczą się w zakresie 20-300 nm.

Ryż. 12. Różnorodność cząstek wirusowych.
Niektórzy przedstawiciele osiągają długość 1-1,5 mikrona. Budowa wirusa polega na otoczeniu materiału genetycznego specjalną ramką białkową (kapsydem), charakteryzującą się różnorodnymi kształtami (spiralny, ikozaedryczny, kulisty). Niektóre wirusy mają również otoczkę na górze utworzoną z błony komórkowej gospodarza (superkapsyd). Na przykład (ryc. 13) jest czynnikiem wywołującym chorobę zwaną AIDS. Zawiera RNA jako materiał genetyczny i wpływa na pewien typ komórek układu odpornościowego (pomocnicze limfocyty T).

Ryż. 13. Struktura ludzkiego wirusa niedoboru odporności.
Bakterie to najstarsza grupa organizmów występująca obecnie na Ziemi. Pierwsze bakterie pojawiły się prawdopodobnie ponad 3,5 miliarda lat temu i przez prawie miliard lat były jedynymi żywymi istotami na naszej planecie. Ponieważ byli to pierwsi przedstawiciele żywej natury, ich ciało miało prymitywną strukturę.
Z biegiem czasu ich budowa stawała się coraz bardziej złożona, jednak do dziś bakterie uważane są za najbardziej prymitywne organizmy jednokomórkowe. Co ciekawe, niektóre bakterie nadal zachowują prymitywne cechy swoich starożytnych przodków. Obserwuje się to u bakterii żyjących w gorących źródłach siarkowych i beztlenowym błocie na dnie zbiorników.
Większość bakterii jest bezbarwna. Tylko nieliczne są fioletowe lub zielone. Ale kolonie wielu bakterii mają jasny kolor, co jest spowodowane uwolnieniem kolorowej substancji do środowiska lub pigmentacją komórek.
Odkrywcą świata bakterii był Antony Leeuwenhoek, holenderski przyrodnik z XVII wieku, który jako pierwszy stworzył doskonały mikroskop powiększający, powiększający przedmioty 160-270 razy.
Bakterie są klasyfikowane jako prokarioty i zaliczane do odrębnego królestwa – bakterii.
Figura
Bakterie to liczne i różnorodne organizmy. Różnią się kształtem.
| Nazwa bakterii | Kształt bakterii | Obraz bakterii |
| Cocci | W kształcie kuli | |
| Bakcyl |  | W kształcie pręta |
| Wibracja | W kształcie przecinka | |
| Spirylla |  | Spirala |
| Streptokoki |  | Łańcuch ziarniaków |
| Gronkowiec |  | Skupiska ziarniaków |
| Diplokok | Dwie okrągłe bakterie zamknięte w jednej torebce śluzowej |
Metody transportu
Wśród bakterii wyróżnia się formy mobilne i nieruchome. Ruchy poruszają się w wyniku skurczów przypominających fale lub za pomocą wici (skręconych spiralnych nici), które składają się ze specjalnego białka zwanego flageliną. Może występować jedna lub więcej wici. U niektórych bakterii znajdują się one na jednym końcu komórki, u innych na dwóch lub na całej powierzchni.

Ale ruch jest także nieodłączną cechą wielu innych bakterii, którym brakuje wici. W ten sposób bakterie pokryte na zewnątrz śluzem są zdolne do ruchu ślizgowego.
Niektóre bakterie wodne i glebowe pozbawione wici mają wakuole gazowe w cytoplazmie. W komórce może znajdować się 40-60 wakuoli. Każdy z nich wypełniony jest gazem (prawdopodobnie azotem). Regulując ilość gazu w wakuolach, bakterie wodne mogą zanurzyć się w słupie wody lub wydostać się na jego powierzchnię, a bakterie glebowe mogą przemieszczać się w kapilarach glebowych.
Siedlisko
Ze względu na prostotę organizacji i bezpretensjonalność bakterie są szeroko rozpowszechnione w przyrodzie. Bakterie można znaleźć wszędzie: w kropli nawet najczystszej wody źródlanej, w ziarnach gleby, w powietrzu, na skałach, w polarnym śniegu, piaskach pustyni, na dnie oceanu, w ropie wydobywanej z dużych głębokości, a nawet w woda z gorących źródeł o temperaturze około 80°C. Żyją na roślinach, owocach, różnych zwierzętach oraz u ludzi w jelitach, jamie ustnej, kończynach i na powierzchni ciała.
Bakterie są najmniejszymi i najliczniejszymi żywymi stworzeniami. Dzięki swoim niewielkim rozmiarom z łatwością wnikają we wszelkie pęknięcia, szczeliny czy pory. Bardzo wytrzymały i przystosowany do różnych warunków życia. Tolerują suszenie, ekstremalne zimno i ogrzewanie do 90°C, nie tracąc przy tym swojej żywotności.
Praktycznie nie ma miejsca na Ziemi, gdzie nie występują bakterie, choć w zróżnicowanych ilościach. Warunki życia bakterii są zróżnicowane. Niektóre z nich wymagają tlenu atmosferycznego, inne go nie potrzebują i są w stanie żyć w środowisku beztlenowym.
W powietrzu: bakterie przedostają się do górnych warstw atmosfery na odległość do 30 km. i więcej.
Szczególnie dużo jest ich w glebie. 1 g gleby może zawierać setki milionów bakterii.
W wodzie: w powierzchniowych warstwach wody w zbiornikach otwartych. Pożyteczne bakterie wodne mineralizują pozostałości organiczne.
W organizmach żywych: bakterie chorobotwórcze dostają się do organizmu ze środowiska zewnętrznego, ale tylko w sprzyjających warunkach powodują choroby. Symbiotyki żyją w narządach trawiennych, pomagając rozkładać i wchłaniać pokarm oraz syntetyzować witaminy.
Struktura zewnętrzna
Komórka bakteryjna pokryta jest specjalną gęstą otoczką – ścianą komórkową, która pełni funkcje ochronne i podporowe, a także nadaje bakterii trwały, charakterystyczny kształt. Ściana komórkowa bakterii przypomina ścianę komórki roślinnej. Jest przepuszczalny: przez niego składniki odżywcze swobodnie przedostają się do komórki, a produkty przemiany materii wychodzą do środowiska. Często bakterie wytwarzają dodatkową warstwę ochronną śluzu na ścianie komórkowej - kapsułkę. Grubość kapsułki może być wielokrotnie większa niż średnica samej komórki, ale może być również bardzo mała. Kapsuła nie jest istotną częścią komórki, powstaje w zależności od warunków, w jakich znajdują się bakterie. Chroni bakterie przed wysychaniem.
Na powierzchni niektórych bakterii występują długie wici (jedna, dwie lub wiele) lub krótkie, cienkie kosmki. Długość wici może być wielokrotnie większa niż wielkość ciała bakterii. Bakterie poruszają się za pomocą wici i kosmków.

Struktura wewnętrzna
Wewnątrz komórki bakteryjnej znajduje się gęsta, nieruchoma cytoplazma. Ma strukturę warstwową, nie ma wakuoli, dlatego w samej substancji cytoplazmy znajdują się różne białka (enzymy) i rezerwowe składniki odżywcze. Komórki bakteryjne nie mają jądra. Substancja niosąca informację dziedziczną koncentruje się w centralnej części ich komórki. Bakterie, - kwas nukleinowy - DNA. Ale ta substancja nie tworzy się w jądrze.
Wewnętrzna organizacja komórki bakteryjnej jest złożona i ma swoje specyficzne cechy. Cytoplazma jest oddzielona od ściany komórkowej błoną cytoplazmatyczną. W cytoplazmie znajduje się główna substancja lub macierz, rybosomy i niewielka liczba struktur błonowych, które pełnią różnorodne funkcje (analogi mitochondriów, retikulum endoplazmatycznego, aparatu Golgiego). Cytoplazma komórek bakteryjnych często zawiera granulki o różnych kształtach i rozmiarach. Granulki mogą składać się ze związków, które służą jako źródło energii i węgla. Krople tłuszczu znajdują się także w komórce bakteryjnej.
W centralnej części komórki zlokalizowana jest substancja jądrowa - DNA, która nie jest oddzielona od cytoplazmy błoną. Jest to analog jądra - nukleoid. Nukleoid nie ma błony, jąderka ani zestawu chromosomów.
Metody jedzenia
Bakterie mają różne metody żywienia. Wśród nich są autotrofy i heterotrofy. Autotrofy to organizmy zdolne do samodzielnego wytwarzania substancji organicznych do odżywiania.
Rośliny potrzebują azotu, ale same nie mogą pobierać azotu z powietrza. Niektóre bakterie łączą cząsteczki azotu w powietrzu z innymi cząsteczkami, w wyniku czego powstają substancje dostępne dla roślin.
Bakterie te osadzają się w komórkach młodych korzeni, co prowadzi do powstawania zgrubień na korzeniach, zwanych guzkami. Takie guzki tworzą się na korzeniach roślin z rodziny strączkowych i niektórych innych roślin.
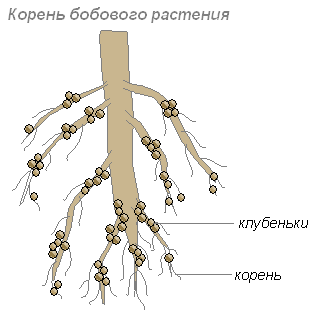
Korzenie dostarczają bakteriom węglowodanów, a bakterie korzeniom substancji zawierających azot, które mogą zostać wchłonięte przez roślinę. Ich wspólne pożycie przynosi obopólne korzyści.
Korzenie roślin wydzielają wiele substancji organicznych (cukrów, aminokwasów i innych), którymi żywią się bakterie. Dlatego szczególnie wiele bakterii osadza się w warstwie gleby otaczającej korzenie. Bakterie te przekształcają martwe resztki roślin w substancje dostępne dla roślin. Ta warstwa gleby nazywa się ryzosferą.
Istnieje kilka hipotez dotyczących przenikania bakterii guzkowych do tkanki korzenia:
- poprzez uszkodzenie tkanki naskórka i kory;
- przez włośniki;
- tylko przez młodą błonę komórkową;
- dzięki bakteriom towarzyszącym wytwarzającym enzymy pektynolityczne;
- poprzez stymulację syntezy kwasu B-indolooctowego z tryptofanu, zawsze obecnego w wydzielinach korzeni roślin.
Proces wprowadzania bakterii guzkowych do tkanki korzenia składa się z dwóch faz:
- infekcja włośników;
- proces powstawania guzków.
W większości przypadków atakująca komórka aktywnie się namnaża, tworzy tzw. nici infekcyjne i w postaci takich nici przedostaje się do tkanki roślinnej. Bakterie guzkowe wychodzące z nici infekcyjnej nadal namnażają się w tkance żywiciela.

Komórki roślinne wypełnione szybko namnażającymi się komórkami bakterii guzkowych zaczynają się szybko dzielić. Połączenie młodego guzka z korzeniem rośliny strączkowej odbywa się dzięki wiązkom naczyniowo-włóknistym. W okresie funkcjonowania guzki są zwykle gęste. Do czasu osiągnięcia optymalnej aktywności guzki przybierają różowy kolor (dzięki pigmentowi leghemoglobiny). Tylko te bakterie, które zawierają leghemoglobinę, są zdolne do wiązania azotu.
Bakterie brodawkowe wytwarzają dziesiątki i setki kilogramów nawozu azotowego na hektar gleby.
Metabolizm
Bakterie różnią się między sobą metabolizmem. U niektórych zachodzi przy udziale tlenu, u innych – bez niego.
Większość bakterii żywi się gotowymi substancjami organicznymi. Tylko nieliczne z nich (niebiesko-zielone lub sinice) są zdolne do tworzenia substancji organicznych z nieorganicznych. Odegrały ważną rolę w akumulacji tlenu w atmosferze ziemskiej.
Bakterie absorbują substancje z zewnątrz, rozrywają swoje cząsteczki na kawałki, składają z tych części otoczkę i uzupełniają ich zawartość (w ten sposób rosną) i wyrzucają niepotrzebne cząsteczki. Otoczka i błona bakterii pozwalają jej wchłonąć tylko niezbędne substancje.

Gdyby otoczka i błona bakterii były całkowicie nieprzepuszczalne, do komórki nie dostałyby się żadne substancje. Gdyby były przepuszczalne dla wszystkich substancji, zawartość komórki mieszałaby się z pożywką – roztworem, w którym żyje bakteria. Aby przetrwać, bakterie potrzebują otoczki, która umożliwia przedostanie się niezbędnych substancji, ale nie substancji niepotrzebnych.
Bakteria wchłania znajdujące się w jej pobliżu składniki odżywcze. Co się potem dzieje? Jeśli potrafi poruszać się samodzielnie (poruszając wicią lub odpychając śluz), to porusza się, aż znajdzie potrzebne substancje.
Jeśli nie może się poruszać, czeka, aż dyfuzja (zdolność cząsteczek jednej substancji do wnikania w gąszcz cząsteczek innej substancji) sprowadzi do niego niezbędne cząsteczki.
Bakterie wraz z innymi grupami mikroorganizmów wykonują ogromną pracę chemiczną. Przekształcając różne związki, otrzymują energię i składniki odżywcze niezbędne do życia. Procesy metaboliczne, sposoby pozyskiwania energii i zapotrzebowanie na materiały do budowy substancji ich ciał są u bakterii zróżnicowane.
Inne bakterie zaspokajają całe swoje zapotrzebowanie na węgiel niezbędny do syntezy substancji organicznych w organizmie kosztem związków nieorganicznych. Nazywa się je autotrofami. Bakterie autotroficzne potrafią syntetyzować substancje organiczne z nieorganicznych. Wśród nich są:
Chemosynteza
Wykorzystanie energii promienistej jest najważniejszym, ale nie jedynym sposobem wytworzenia materii organicznej z dwutlenku węgla i wody. Wiadomo, że bakterie do takiej syntezy jako źródło energii wykorzystują nie światło słoneczne, ale energię wiązań chemicznych zachodzących w komórkach organizmów podczas utleniania niektórych związków nieorganicznych - siarkowodoru, siarki, amoniaku, wodoru, kwasu azotowego, związków żelaza żelazo i mangan. Wykorzystują materię organiczną powstałą przy użyciu tej energii chemicznej do budowy komórek swojego ciała. Dlatego proces ten nazywa się chemosyntezą.
Najważniejszą grupą mikroorganizmów chemosyntetycznych są bakterie nitryfikacyjne. Bakterie te żyją w glebie i utleniają amoniak powstający podczas rozkładu pozostałości organicznych do kwasu azotowego. Ten ostatni reaguje ze związkami mineralnymi gleby, zamieniając się w sole kwasu azotowego. Proces ten odbywa się w dwóch fazach.
Bakterie żelazne przekształcają żelazo żelazne w żelazo tlenkowe. Powstały wodorotlenek żelaza osadza się i tworzy tzw. rudę darniową.
Niektóre mikroorganizmy istnieją w wyniku utleniania wodoru cząsteczkowego, zapewniając w ten sposób autotroficzną metodę odżywiania.
Charakterystyczną cechą bakterii wodorowych jest zdolność do przejścia na heterotroficzny tryb życia, gdy zapewnione są im związki organiczne i brak wodoru.
Zatem chemoautotrofy są typowymi autotrofami, ponieważ niezależnie syntetyzują niezbędne związki organiczne z substancji nieorganicznych i nie pobierają ich w postaci gotowej od innych organizmów, takich jak heterotrofy. Bakterie chemoautotroficzne różnią się od roślin fototroficznych całkowitą niezależnością od światła jako źródła energii.
Fotosynteza bakteryjna
Niektóre bakterie siarkowe zawierające pigmenty (fioletowe, zielone), zawierające specyficzne pigmenty - bakteriochlorofile, są w stanie absorbować energię słoneczną, za pomocą której siarkowodór w ich ciałach rozkłada się i uwalnia atomy wodoru w celu przywrócenia odpowiednich związków. Proces ten ma wiele wspólnego z fotosyntezą, a różni się jedynie tym, że u bakterii fioletowych i zielonych donorem wodoru jest siarkowodór (czasami kwasy karboksylowe), a u roślin zielonych jest to woda. W obu przypadkach separacja i transfer wodoru odbywa się dzięki energii pochłoniętych promieni słonecznych.
Ta fotosynteza bakteryjna, która zachodzi bez uwalniania tlenu, nazywa się fotoredukcją. Fotoredukcja dwutlenku węgla wiąże się z przeniesieniem wodoru nie z wody, ale z siarkowodoru:
6СО 2 +12Н 2 S+hv → С6Н 12 О 6 +12S=6Н 2 О
Biologiczne znaczenie chemosyntezy i fotosyntezy bakteryjnej w skali planetarnej jest stosunkowo niewielkie. Jedynie bakterie chemosyntetyzujące odgrywają znaczącą rolę w procesie obiegu siarki w przyrodzie. Wchłaniana przez rośliny zielone w postaci soli kwasu siarkowego, siarka ulega redukcji i staje się częścią cząsteczek białka. Co więcej, gdy martwe szczątki roślin i zwierząt zostaną zniszczone przez bakterie gnilne, siarka uwalnia się w postaci siarkowodoru, który jest utleniany przez bakterie siarkowe do wolnej siarki (lub kwasu siarkowego), tworząc w glebie siarczyny dostępne dla roślin. Bakterie chemo- i fotoautotroficzne są niezbędne w cyklu azotu i siarki.
Sporulacja
Zarodniki tworzą się wewnątrz komórki bakteryjnej. W procesie sporulacji komórka bakteryjna przechodzi szereg procesów biochemicznych. Zmniejsza się w nim ilość wolnej wody i zmniejsza się aktywność enzymatyczna. Zapewnia to odporność zarodników na niekorzystne warunki środowiskowe (wysoka temperatura, wysokie stężenie soli, suszenie itp.). Sporulacja jest charakterystyczna tylko dla niewielkiej grupy bakterii.

Zarodniki są opcjonalnym etapem cyklu życiowego bakterii. Sporulacja rozpoczyna się dopiero w przypadku braku składników odżywczych lub gromadzenia się produktów przemiany materii. Bakterie w postaci zarodników mogą pozostawać w stanie uśpienia przez długi czas. Zarodniki bakterii mogą wytrzymać długotrwałe gotowanie i bardzo długie zamrażanie. Gdy zaistnieją sprzyjające warunki, zarodnik kiełkuje i staje się żywotny. Zarodniki bakterii są przystosowaniem do przetrwania w niesprzyjających warunkach.
Reprodukcja
Bakterie rozmnażają się, dzieląc jedną komórkę na dwie. Po osiągnięciu określonej wielkości bakteria dzieli się na dwie identyczne bakterie. Następnie każdy z nich zaczyna żerować, rośnie, dzieli się i tak dalej.

Po wydłużeniu komórek stopniowo tworzy się przegroda poprzeczna, a następnie komórki potomne oddzielają się; U wielu bakterii w określonych warunkach komórki po podziale pozostają połączone w charakterystyczne grupy. W tym przypadku w zależności od kierunku płaszczyzny podziału i liczby podziałów powstają różne kształty. Rozmnażanie przez pączkowanie występuje u bakterii jako wyjątek.
W sprzyjających warunkach podział komórek u wielu bakterii następuje co 20-30 minut. Przy tak szybkiej reprodukcji potomstwo jednej bakterii w ciągu 5 dni może utworzyć masę, która może wypełnić wszystkie morza i oceany. Z prostych obliczeń wynika, że dziennie mogą powstać 72 pokolenia (720 000 000 000 000 000 000 komórek). W przeliczeniu na masę - 4720 ton. Jednak w naturze tak się nie dzieje, gdyż większość bakterii szybko ginie pod wpływem światła słonecznego, suszenia, braku pożywienia, ogrzewania do 65-100°C, w wyniku walki między gatunkami itp.
Bakteria (1) po wchłonięciu wystarczającej ilości pożywienia zwiększa swój rozmiar (2) i zaczyna przygotowywać się do rozmnażania (podziału komórki). Jej DNA (w bakterii cząsteczka DNA jest zamknięta w pierścieniu) podwaja się (bakteria wytwarza kopię tej cząsteczki). Obie cząsteczki DNA (3,4) przyczepiają się do ściany bakterii i w miarę wydłużania się bakterii oddalają się od nich (5,6). Najpierw dzieli się nukleotyd, potem cytoplazma.

Po rozbieżności dwóch cząsteczek DNA na bakterii pojawia się zwężenie, które stopniowo dzieli organizm bakterii na dwie części, z których każda zawiera cząsteczkę DNA (7).
Zdarza się (u Bacillus subtilis), że dwie bakterie sklejają się ze sobą i tworzy się pomiędzy nimi most (1,2).

Skoczek transportuje DNA z jednej bakterii do drugiej (3). W jednej bakterii cząsteczki DNA przeplatają się, w niektórych miejscach sklejają (4), a następnie wymieniają odcinki (5).
Rola bakterii w przyrodzie
wir
Bakterie są najważniejszym ogniwem w ogólnym cyklu substancji w przyrodzie. Rośliny tworzą złożone substancje organiczne z dwutlenku węgla, wody i soli mineralnych znajdujących się w glebie. Substancje te wracają do gleby wraz z martwymi grzybami, roślinami i zwłokami zwierząt. Bakterie rozkładają złożone substancje na proste, które następnie są wykorzystywane przez rośliny.
Bakterie niszczą złożone substancje organiczne martwych roślin i zwłok zwierząt, wydaliny organizmów żywych i różne odpady. Żywiąc się tymi substancjami organicznymi, saprofityczne bakterie rozkładu zamieniają je w humus. To swego rodzaju sanitariusze naszej planety. W ten sposób bakterie aktywnie uczestniczą w cyklu substancji w przyrodzie.
Tworzenie gleby
Ponieważ bakterie są rozmieszczone niemal wszędzie i występują w ogromnych ilościach, w dużej mierze determinują różnorodne procesy zachodzące w przyrodzie. Jesienią opadają liście drzew i krzewów, zamierają nadziemne pędy traw, opadają stare gałęzie, a od czasu do czasu opadają pnie starych drzew. Wszystko to stopniowo zamienia się w humus. W 1 cm3. Powierzchniowa warstwa gleby leśnej zawiera setki milionów saprofitycznych bakterii glebowych kilku gatunków. Bakterie te przekształcają próchnicę w różne minerały, które mogą być wchłaniane z gleby przez korzenie roślin.
Niektóre bakterie glebowe potrafią pobierać azot z powietrza, wykorzystując go w procesach życiowych. Te bakterie wiążące azot żyją niezależnie lub osiedlają się w korzeniach roślin strączkowych. Bakterie te po wniknięciu do korzeni roślin strączkowych powodują wzrost komórek korzeni i powstawanie na nich guzków.
Bakterie te wytwarzają związki azotu wykorzystywane przez rośliny. Bakterie pozyskują z roślin węglowodany i sole mineralne. Zatem istnieje ścisły związek między rośliną strączkową a bakteriami brodawkowymi, co jest korzystne zarówno dla jednego, jak i drugiego organizmu. Zjawisko to nazywa się symbiozą.
Dzięki symbiozie z bakteriami brodawkowymi rośliny strączkowe wzbogacają glebę w azot, przyczyniając się do zwiększenia plonów.
Dystrybucja w przyrodzie
Mikroorganizmy są wszechobecne. Jedynymi wyjątkami są kratery aktywnych wulkanów i małe obszary w epicentrach eksplodowanych bomb atomowych. Ani niskie temperatury Antarktydy, ani wrzące strumienie gejzerów, ani nasycone roztwory soli w basenach solnych, ani silne nasłonecznienie szczytów górskich, ani ostre napromieniowanie reaktorów jądrowych nie zakłócają istnienia i rozwoju mikroflory. Wszystkie żywe istoty stale wchodzą w interakcję z mikroorganizmami, często będąc nie tylko ich magazynami, ale także ich dystrybutorami. Mikroorganizmy są mieszkańcami naszej planety i aktywnie eksplorują najbardziej niesamowite naturalne substraty.
Mikroflora gleby
Liczba bakterii w glebie jest niezwykle duża – setki milionów i miliardów osobników na gram. W glebie jest ich znacznie więcej niż w wodzie i powietrzu. Zmienia się całkowita liczba bakterii w glebie. Liczba bakterii zależy od rodzaju gleby, jej stanu i głębokości warstw.
Na powierzchni cząstek gleby mikroorganizmy gromadzą się w małych mikrokoloniach (po 20–100 komórek). Często rozwijają się w grubości skrzepów materii organicznej, na żywych i obumierających korzeniach roślin, w cienkich kapilarach i wewnątrz grudek.
Mikroflora glebowa jest bardzo zróżnicowana. Występują tu różne fizjologiczne grupy bakterii: bakterie gnilne, bakterie nitryfikacyjne, bakterie wiążące azot, bakterie siarkowe itp. Wśród nich są bakterie tlenowe i beztlenowe, formy zarodnikowe i niezarodnikowe. Mikroflora jest jednym z czynników tworzących glebę.
Obszarem rozwoju mikroorganizmów w glebie jest strefa sąsiadująca z korzeniami żywych roślin. Nazywa się to ryzosferą, a całość zawartych w niej mikroorganizmów nazywa się mikroflorą ryzosfery.
Mikroflora zbiorników wodnych
Woda jest naturalnym środowiskiem, w którym rozwijają się licznie mikroorganizmy. Większość z nich przedostaje się do wody z gleby. Czynnik decydujący o liczbie bakterii w wodzie i obecności w niej składników odżywczych. Najczystsza woda pochodzi ze studni artezyjskich i źródeł. Otwarte zbiorniki i rzeki są bardzo bogate w bakterie. Najwięcej bakterii występuje w powierzchniowych warstwach wody, bliżej brzegu. W miarę oddalania się od brzegu i zwiększania głębokości liczba bakterii maleje.
Czysta woda zawiera 100-200 bakterii na ml, a zanieczyszczona woda zawiera 100-300 tysięcy i więcej. W osadzie dennym znajduje się wiele bakterii, zwłaszcza w warstwie powierzchniowej, gdzie bakterie tworzą film. Film ten zawiera dużo bakterii siarkowych i żelaznych, które utleniają siarkowodór do kwasu siarkowego, zapobiegając w ten sposób umieraniu ryb. W mule występuje więcej form zarodnikotwórczych, natomiast w wodzie dominują formy niezarodnikowe.
Mikroflora wodna pod względem składu gatunkowego jest podobna do mikroflory glebowej, ale występują też formy specyficzne. Niszcząc różne odpady dostające się do wody, mikroorganizmy stopniowo dokonują tzw. biologicznego oczyszczania wody.
Mikroflora powietrza
Mikroflora powietrza jest mniej liczna niż mikroflora gleby i wody. Bakterie wraz z pyłem unoszą się w powietrzu, mogą tam pozostać przez pewien czas, a następnie osadzić się na powierzchni ziemi i ginąć z powodu braku pożywienia lub pod wpływem promieni ultrafioletowych. Liczba mikroorganizmów w powietrzu zależy od strefy geograficznej, ukształtowania terenu, pory roku, zanieczyszczenia pyłem itp. Każda cząstka kurzu jest nośnikiem mikroorganizmów. Większość bakterii znajduje się w powietrzu nad przedsiębiorstwami przemysłowymi. Powietrze na obszarach wiejskich jest czystsze. Najczystsze powietrze panuje nad lasami, górami i obszarami zaśnieżonymi. Górne warstwy powietrza zawierają mniej drobnoustrojów. Mikroflora powietrza zawiera wiele bakterii pigmentowanych i zarodnikowych, które są bardziej odporne na działanie promieni ultrafioletowych niż inne.
Mikroflora organizmu człowieka
Organizm ludzki, nawet całkowicie zdrowy, jest zawsze nośnikiem mikroflory. Kiedy organizm ludzki ma kontakt z powietrzem i glebą, na odzieży i skórze osadzają się różne mikroorganizmy, w tym chorobotwórcze (pałeczki tężca, zgorzel gazowa itp.). Zanieczyszczone są najczęściej narażone części ciała człowieka. Na rękach znajdują się E. coli i gronkowce. W jamie ustnej żyje ponad 100 rodzajów drobnoustrojów. Jama ustna ze swoją temperaturą, wilgotnością i pozostałościami składników odżywczych jest doskonałym środowiskiem do rozwoju mikroorganizmów.
Żołądek ma odczyn kwaśny, więc większość znajdujących się w nim mikroorganizmów umiera. Począwszy od jelita cienkiego, odczyn staje się zasadowy, tj. korzystne dla drobnoustrojów. Mikroflora w jelicie grubym jest bardzo zróżnicowana. Każdy dorosły człowiek wydala dziennie w odchodach około 18 miliardów bakterii, tj. więcej osób niż ludzi na świecie.
Narządy wewnętrzne, które nie są połączone ze środowiskiem zewnętrznym (mózg, serce, wątroba, pęcherz moczowy itp.), są zwykle wolne od drobnoustrojów. Drobnoustroje dostają się do tych narządów dopiero w czasie choroby.
Bakterie w obiegu substancji
Mikroorganizmy w ogóle, a bakterie w szczególności odgrywają dużą rolę w biologicznie ważnych cyklach substancji na Ziemi, dokonując przemian chemicznych całkowicie niedostępnych ani dla roślin, ani dla zwierząt. Różne etapy cyklu pierwiastków przeprowadzane są przez organizmy różnych typów. Istnienie poszczególnych grup organizmów zależy od przemian chemicznych pierwiastków przeprowadzanych przez inne grupy.
Cykl azotowy
Cykliczne przemiany związków azotowych odgrywają podstawową rolę w dostarczaniu niezbędnych form azotu organizmom biosfery o różnych potrzebach żywieniowych. Ponad 90% całkowitego wiązania azotu wynika z aktywności metabolicznej niektórych bakterii.
Obieg węgla
Biologiczna przemiana węgla organicznego w dwutlenek węgla, której towarzyszy redukcja tlenu cząsteczkowego, wymaga wspólnej aktywności metabolicznej różnych mikroorganizmów. Wiele bakterii tlenowych przeprowadza całkowite utlenianie substancji organicznych. W warunkach tlenowych związki organiczne są początkowo rozkładane w procesie fermentacji, a organiczne produkty końcowe fermentacji są dalej utleniane w procesie oddychania beztlenowego, jeśli obecne są nieorganiczne akceptory wodoru (azotany, siarczany lub CO2).
Cykl siarkowy
Siarka jest dostępna dla organizmów żywych głównie w postaci rozpuszczalnych siarczanów lub zredukowanych organicznych związków siarki.
Cykl żelaza
Niektóre zbiorniki słodkowodne zawierają wysokie stężenia zredukowanych soli żelaza. W takich miejscach rozwija się specyficzna mikroflora bakteryjna – bakterie żelazowe, które utleniają zredukowane żelazo. Biorą udział w powstawaniu rud darniowych i źródeł wód bogatych w sole żelaza.
Bakterie to najstarsze organizmy, które pojawiły się w Archaiku około 3,5 miliarda lat temu. Przez około 2,5 miliarda lat dominowały na Ziemi, tworząc biosferę i brały udział w tworzeniu atmosfery tlenowej.
Bakterie są jednymi z żywych organizmów o najprostszej strukturze (z wyjątkiem wirusów). Uważa się, że są to pierwsze organizmy, które pojawiły się na Ziemi.
Według nowego systemu wyróżnia się trzy domeny: „Bakterie” (eubakterie), „Archaea” (archaebakterie) i „Eisagua” (eukarioty). Domeny obejmują typy, klasy, rzędy, rodziny, rodzaje i gatunki.
Bakteria są prokariotami (w komórce prokariotycznej jądro, zwane nukleoidem, nie posiada otoczki jądrowej, jąderka i histonów, a cytoplazma nie zawiera wysoce zorganizowanych organelli).
Do grubościennego, Gram-dodatniego bakteria obejmują: formy kuliste lub ziarniaki (gronkowce, paciorkowce, pneumokoki); w kształcie pręta, w tym maczugowców, prątki i bifidobakterie; promieniowce (bakterie rozgałęzione, nitkowate).
Formy kuliste lub ziarniaki - bakterie kuliste o wielkości 0,5-1,0 mikrona; Według względnego ułożenia komórek mikrokoki, diplokoki, paciorkowce, tetrakoki, sarcyna i gronkowce. Mikrokoki (grecki) mikros- mały)
- oddzielnie zlokalizowane komórki lub w formie „pakietów”.
Diplococci (z greckiego.dyplomaci- podwójne) lub sparowane ziarniaki, ułożone parami (pneumokoki, gonokok, meningokoki), ponieważ komórki nie rozdzielają się po podziale. Pneumokoki ma lancetowaty kształt po przeciwnych stronach, oraz gonokok I meningokoki mają kształt ziaren kawy, z wklęsłą powierzchnią zwróconą ku sobie. Streptokoki(z greckiego streptos- łańcuch) - komórki o okrągłym lub wydłużonym kształcie, tworzące łańcuch w wyniku podziału komórek w tej samej płaszczyźnie i utrzymania połączenia między nimi w miejscu podziału. Sarcyny (od łac. sarcyna- kiść, bela) są ułożone w formie „pakietów” po 8 lub więcej ziarniaków, ponieważ powstają podczas podziału komórek w trzech wzajemnie prostopadłych płaszczyznach. Staphylococcus (z greckiego. staphyl- kiść winogron) - ziarniaki powstałe w postaci kiści winogron w wyniku podziału w różnych płaszczyznach.
Bakterie w kształcie pręta różnią się rozmiarem, kształtem końców komórek i względnym położeniem komórek. Długość komórek waha się od 1,0 do 8,0 µm, grubość – od 0,5 do 2,0 µm. Pręciki mogą mieć kształt regularny (Escherichia coli itp.) i nieregularny (maczugowca itp.), w tym rozgałęziony, np. w promieniowce. Lekko zakrzywione pręty nazywane są vibrios (Vibrio cholerae). Większość w kształcie pręta bakteria jest umiejscowiony losowo, gdyż po podziale komórki się rozchodzą. Jeżeli po podziale komórki pozostają połączone wspólnymi fragmentami ściany komórkowej i nie rozchodzą się, wówczas układają się względem siebie pod kątem (Corynebacterium diphtheria) lub tworzą łańcuch (bacillus wąglika).
Ryż. 3.1. RozmazzEscherichia coliIStaphylococcus aureus.Malarstwo Grama
W starym podręczniku Bergey Manual of Systematic Bacteriology bakterie podzielono zgodnie z charakterystyką ściany komórkowej bakterii na 4 działy: Gracilicutes - eubakterie o cienkiej ścianie komórkowej, Gram-ujemne; Firmicutes - eubakterie o grubej ścianie komórkowej, Gram-dodatnie; Tenericutes - eubakterie bez ściany komórkowej; Mendosicutes to archaebakterie z wadliwą ścianą komórkową. Każdy oddział został podzielony na sekcje lub grupy w oparciu o barwienie metodą Grama, kształt komórek, zapotrzebowanie na tlen, ruchliwość, cechy metaboliczne i odżywcze.
Tabela 3.1. Według drugiego wydania (2001) Podręcznika Bergeya bakterie dzielą się na dwie domeny: „Bakteria»
I «
Archeony»
|
Domena "Bakteria"(eubakterie) |
Domena „Archea”(archebakterie) |
|
W domenie «
Bakteria»
Można wyizolować następujące bakterie: |
Archebakterie nie zawierają peptydoglikanu w ścianie komórkowej. Mają specjalne rybosomy i rybosomalny RNA (rRNA). Termin „archaebakteria” pojawił się w 1977 roku. Jest to jedna z najstarszych form życia, co oznacza przedrostek „arche”. Nie ma wśród nich patogenów chorób zakaźnych. |
|
Większość bakterii Gram-ujemnych grupuje się w gromadę Proteobacteria na podstawie podobieństwa w rybosomalnym RNA („Proteobacteria” – od greckiego boga Proteusa, który przybierał różne formy). Wyewoluowały od wspólnego fotosyntetycznego przodka. |
|
Do domeny "Bakteria„obejmuje 22 typy, z których następujące mają znaczenie medyczne (według Bergi, 2001):
TypWIV. Deinococcus-ThermusKlasaI. Deinokoki
Rząd I. Rodzina Deinococcales I. Rodzaj Deinococcaceae1. Deinokoki
TypWXII. Proteobakterie
KlasaI. Alphaproteobakterie Zamówienie II. Rodzina Rickettsiales I. Rickettsiaceae Rodzaj I. Rickettsia Rodzaj I. Orientia Rodzaj III. Rodzina Wolbachia I. Ehrlichiaceae (b rodzajów) Rodzaj I. Ehrlichia Rodzaj I. Rodzina Aegyptianella III. Holosporaceae (8 rodzajów) Rząd VI. Rodzina Rhizobiales I. Bartonellaceae Rodzaj I. Rodzina Bartonella III. Brucellaceae Rodzaj I. Brucella
KlasaII. Betaproteobakterie Rząd I. Rodzina Burkholderiales I. Burkholderiaceae Rodzaj I. Rodzina Burkholderia IV. Alcaligenaceae Rodzaj I. Alcaligenes Rodzaj III. Bordetella Rząd IV. Rodzina Neisseriales I. Neisseriaceae Rodzaj I. Rodzaj Neisseria VI.Eikenella Rodzaj IX.Kingella Rząd V. Rodzina Nitrozomonadales II. Spirillaceae Rodzaj I. Spirillum
KlasaIII. Gammaproteobakterie Zakon V. Rodzina Thiotrichales III. Francisellaceae Rodzaj I. Francisella Rząd VI. Rodzina Legionellales I. Legionellaceae Rodzaj I. Rodzina Legionella II. Coxiellaceae Rodzaj I. Coxiella Rząd IX. Rodzina Pseudomonadales I. Pseudomonadaceae Rodzaj I. Rodzina Pseudomonas I. Moraxellaceae Rodzaj I. Moraxella Rodzaj I. Acinetobacter Rząd XI. Rodzina Vibrionales I. Vibrionaceae Rodzaj I. Vibrio Rząd XII. Rodzina Aeromonadales I. Aeromonadaceae Rodzaj I. Aeromonas Rząd XIII. Rodzina Enterobacteri I. Enterobacteriaceae Rodzaj I. Enterobacter Rodzaj VIII. Calymmatobacterium rodzaj X. Citrobacter rodzaj XI. Edwardsiella Rodzaj XII. Erwinia Rodzaj XIII Escherichia Rodzaj XV. Hafnia
Rodzaj XVI. Klebsiella Rodzaj XVII. Kluyvera Rodzaj XXI. Morganella rodzaj XXVI. Plesiomonas, rodzaj XXVIII. Proteusz, rodzaj XXIX. Providencia RodXXXY. Rodzaj Salmonelli XXXIII. Rodzaj Serratia XXXIV. Rodzaj Shigella XL. Zakon Yersinia IV. Rodzina Pasteurellales I. Pasteurellaceae Rodzaj I. Pasteurella Rodzaj I. Actinobacillus Rodzaj III. Hemofil
KlasaIV. Deltaproteobakterie Zamówienie II. Rodzina Desulfovibrionales I. Desulfovibrionaceae Rodzaj II. Bilofil
KlasaV. Epsilonproteobakterie Rząd I. Rodzina Campylobacteri I. Rodzaj Campylobacteriaceae I. Rodzina Campylobacter II. Helicobacteriaceae Rodzaj I. Helicobacter Rodzaj Il.Wolinella
TypWXIII. Firmicutes (głównysposóbGram-dodatnie)
KlasaI.
Clostridia Rząd I. Rodzina Clostridiales I. Clostridiaiaceae Rodzaj I. Clostridium Rodzaj IX. Rodzina Sarcinach III. Peptostreptococcus Rodzaj I. Rodzina Peptostreptococcus IV. Eubacteriaceae Rodzaj I. Rodzina Eubacterium V. Peptococcaceae Rodzaj I. Rodzina Peptococcus VII. Acidaminococcaceae Rodzaj XIV. Veillonella KlasaII. Mollicutes Rząd I. Rodzina Mycoplasmatales I. Mycoplasmataceae Rodzaj I. Mycoplasma Rodzaj IV. Ureaplazma KlasaIII. Pałeczki Rząd I. Rodzina Bacillalles I. Bacillaceae Rodzaj I. Rodzina Bacillus I. Planococcaceae Rodzaj I. Planococcus Rodzaj IV Rodzina Sporosarcina IV. Listeriaceae Rodzaj I. Rodzina Listeria V. Staphylococcaceae Rodzaj I. Staphylococcus Rodzaj II. Rząd Gemella I. Rodzina Lactobacillus I. Rodzaj Lactobacillus I. Rodzaj Lactobacillus III. Rodzina Pediococcus II. Aegococcusae Rodzaj I. Rodzina Aerococcus IV. Enterococcaceae Rodzaj I. Enterococcus
Rodzina V. Leuconostocaceae Rodzaj I. Rodzina Leuconostoc VI. Streptococcus rodzaj I. Streptococcus rodzaj II. Lactococcus TypWXIV. ActinobakterieKlasaI. Actinobacteria Podklasa V. Actinomycetales Rząd I. Actinomycetales Podrząd V. Rodzina Actinomycineae I. Actinomycetaceae Rodzaj I. Rodzaj Actinomyces I. Rodzaj Actinobacilum III. Arcanodakterium Rodzaj IV. Podrząd Mobiluncus VI. Rodzina Micrococcineae I. Micrococcaceae Rodzaj I. Micrococcus Rodzaj VI. Rodzaj Rothia VII. Podrząd Stomatococcus VII. Rodzina Corynebacterineae I. Corynebacteriaceae Rodzaj I. Rodzina Corynebacterium I. Mycobacteriaceae Rodzaj IV. Rodzina Mycobacterium V. Nocardiaceae Rodzaj I. Nokardia Rodzaj I. Rhodococcus Podrząd VII. Rodzina Propionibacterineae I. Propionibacteriaceae Rodzaj I. Propionibacterium Rodzina I. Nocardiaceae Rodzaj I. Nokardioides Zamówienie II. Bifidobakterie
Rodzina I. Bifidobacteriaceae Rodzaj I. Bifidobacterium Rodzaj III. Gardnerella TypWXVI. ChlamydieKlasaI. Chlamydie Rząd I. Rodzina Chlamydiales I. Chlamydiaceae Rodzaj I. Chlamydia Rodzaj II. Chlamydofilia TypWXVII. KrętkiKlasaI. Krętki Rząd I. Rodzina Spirochaetales I. Spirochaetaceae Rodzaj I. Rodzaj Spirochaeta I. Borrelia Rodzaj IX. Rodzina Treponemów III. Leptospiraceae Rodzaj I. Leptospira TypWXX. BakterioidetyKlasaI. Bacteroidetes Rząd I. Rodzina Bacteroides I. Bacteroidaceae Rodzaj I. Rodzina Bacteroides III. Porphyromonadaceae Rodzaj I. Rodzina Porphyromonas IV. Prevotellaceae Rodzaj I. Prevotella KlasaII. Flawobakterie Rząd I. Rodzina Flavobacteri I. Flavobacteriaceae Rodzaj I. Flavobacterium

*Lokalizacja zarodników: 1 - centralna, 2 - podterminalna, 3 - końcowa.
Ryż. 3.2.
Chlamydia należą do obligatoryjnych wewnątrzkomórkowych kokoidowych bakterii Gram-ujemnych (czasami Gram-zmiennych). Rozmnażają się wyłącznie w żywych komórkach. Na zewnątrz komórek chlamydie mają kształt kulisty (0,3 µm), są nieaktywne metabolicznie i nazywane są ciałami elementarnymi. Ściana komórkowa ciał elementarnych zawiera główne białko błony zewnętrznej i białko zawierające dużą ilość cysteiny. Ciała elementarne dostają się do komórki nabłonkowej poprzez endocytozę z utworzeniem wakuoli wewnątrzkomórkowej. Wewnątrz komórek powiększają się i przekształcają w dzielące się ciała siatkowe, tworząc skupiska w wakuolach (wtręty). Ciała elementarne powstają z ciał siatkowych, które opuszczają komórki w wyniku egzocytozy lub lizy komórek. Ciała elementarne uwolnione z komórki wchodzą w nowy cykl, infekując inne komórki. U ludzi chlamydia powoduje uszkodzenie oczu, dróg moczowo-płciowych, płuc itp.
Mykoplazma - małe bakterie (0,15-1,0 µm), otoczone błoną cytoplazmatyczną i pozbawione ściany komórkowej. Ze względu na brak ściany komórkowej mykoplazmy są wrażliwe osmotycznie. Mają różne kształty: kokosowy, nitkowaty, w kształcie kolby; podobny do kształtów L (ryc. 3.6). Formy te są widoczne pod mikroskopem z kontrastem fazowym czystych kultur mykoplazm. Mykoplazmy chorobotwórcze powodują przewlekłe infekcje - mykoplazmozę.
Promieniowce - bakterie Gram-dodatnie rozgałęzione, nitkowate lub w kształcie pałeczki. Jego nazwa (z greckiego. akt- Ray, mykes- grzyb) otrzymali w wyniku tworzenia się w dotkniętych tkankach druz - granulek ściśle splecionych nici w postaci promieni rozciągających się od środka i kończących się zgrubieniami w kształcie kolby. Promieniowce mogą dzielić się poprzez fragmentację grzybni na komórki podobne do bakterii pałeczek i bakterii kokosowych. Na strzępkach powietrznych promieniowców mogą tworzyć się zarodniki, które służą do rozmnażania. Zarodniki promieniowca zwykle nie są odporne na ciepło.

Miejsce przyczepu (analogicznie do blefaroplasty)
Ryż. 3.3.Wzór dyfrakcji elektronów fragmentu komórki Treponema pallidum (kontrast ujemny). Według N. M. Ovchinnikova, V. V. Delectorsky'ego
Wspólną gałąź filogenetyczną z promieniowcami tworzą tzw. promieniowce nocardiform (nocardioform) – zbiorowa grupa bakterii o kształcie pręcików i nieregularnych kształtach. Ich poszczególni przedstawiciele tworzą rozgałęzione formy. Należą do nich bakterie z rodzaju Corynebakteria,
Mykobakterie,
Nokardia itp. Promieniowce Nocardi-podobne wyróżniają się obecnością w ścianie komórkowej cukrów arabinozy, galaktozy, a także kwasów mykolowych i dużych ilości kwasów tłuszczowych. Kwasy mikolowe i lipidy ścian komórkowych decydują o kwasoodporności bakterii, w szczególności prątków gruźlicy i trądu (po zabarwieniu według Ziehl-Neelsena są czerwone, a niekwasoodporne bakterie i elementy tkanek, plwocina mają kolor niebieski).
Formy zwinięte to bakterie o kształcie spiralnym, na przykład spirilla, które mają wygląd skręconych komórek w kształcie korkociągu. Spirilla chorobotwórcza obejmuje czynnik sprawczy sodoku (choroba ugryzienia szczura). Do zawiłych należą także Campylobacter, Helicobacter, które mają krzywizny przypominające skrzydło latającej mewy; blisko nich znajdują się także bakterie takie jak krętki. Krętki to cienkie, długie, skręcone (spiralne) bakterie, które różnią się od spirillum ruchliwością ze względu na zmiany zginania komórek. Krętki mają zewnętrzną ścianę komórkową otaczającą protoplazmatyczny cylinder z błoną cytoplazmatyczną. Pod zewnętrzną błoną ściany komórkowej (w peryplazmie) znajdują się włókienka peryplazmatyczne (wici), które jakby owijając się wokół protoplazmatycznego cylindra krętka, nadają mu spiralny kształt (pierwotne loki krętka). Włókna są przyczepione do końców komórki (ryc. 3.3) i są skierowane ku sobie. Drugi koniec włókienek jest wolny. Liczba i rozmieszczenie włókienek różni się w zależności od gatunku. Włókna biorą udział w ruchu krętków, nadając komórkom ruch obrotowy, zginający i translacyjny. W tym przypadku krętki tworzą pętle, loki i zagięcia, które nazywane są lokami wtórnymi.
Krętki nie akceptują dobrze barwników. Barwi się je metodą Romanowskiego-Giemsy lub srebrzeniem i bada w postaci żywej za pomocą mikroskopii z kontrastem fazowym lub w ciemnym polu. Krętki reprezentowane są przez 3 rodzaje chorobotwórcze dla człowieka: Treponema, Borrelia, Leptospira.
Treponema(rodzaj Treponema) mają wygląd cienkich, skręconych korkociągami nitek z 8-12 jednolitymi małymi lokami. Włókna znajdują się wokół protoplastu krętka. Przedstawicielami patogennymi są T. blady- czynnik sprawczy kiły, T. trwałe- czynnik sprawczy choroby tropikalnej ziewa.
Borrelia(rodzaj Borrelia) są dłuższe, mają 3-8 dużych okółków i 8-20 włókienek. Należą do nich czynnik wywołujący nawracającą gorączkę (B. nawracające) i patogeny boreliozy (V. burgdorfery itd.).
Leptospira(rodzaj Leptospira) mają płytkie i częste loki - w postaci skręconej liny. Końce tych krętków są zakrzywione jak haczyki ze zgrubieniami na końcach. Tworząc wtórne loki, przyjmują wygląd liter „S” lub „C”; mają 2 gwinty osiowe. Przedstawiciel patogenny L
