طبقه بندی باکتری ها بر اساس محیط آنها اصول طبقه بندی میکروارگانیسم ها
2.1. سیستماتیک و نامگذاری میکروب ها
دنیای میکروبی را می توان به دو شکل سلولی و غیر سلولی تقسیم کرد. اشکال سلولی میکروب ها توسط باکتری ها، قارچ ها و تک یاخته ها نشان داده می شوند. آنها را می توان میکروارگانیسم نامید. اشکال غیر سلولی توسط ویروس ها، ویروئیدها و پریون ها نشان داده می شوند.
طبقه بندی جدید میکروب های سلولی شامل واحدهای طبقه بندی زیر است: دامنه ها، پادشاهی ها، انواع، طبقات، راسته ها، خانواده ها، جنس ها، گونه ها. طبقه بندی میکروارگانیسم ها بر اساس رابطه ژنتیکی آنها و همچنین ویژگی های مورفولوژیکی، فیزیولوژیکی، آنتی ژنی و بیولوژیکی مولکولی است.
ویروس ها اغلب نه به عنوان ارگانیسم ها، بلکه به عنوان ساختارهای ژنتیکی مستقل در نظر گرفته می شوند، بنابراین آنها به طور جداگانه در نظر گرفته می شوند.
اشکال سلولی میکروب ها به سه حوزه تقسیم می شوند. دامنه ها باکتری هاو آرکی باکتری هاشامل میکروب هایی با ساختار سلولی نوع پروکاریوتی است. نمایندگان دامنه یوکاریایوکاریوت هستند از 4 پادشاهی تشکیل شده است:
پادشاهی قارچ (قارچ، Eumycota)؛
پادشاهی تک یاخته ها (پرتوزوآر)؛
پادشاهی ها کرومیستا(صفحات کرومی)؛
میکروب هایی با موقعیت طبقه بندی نامشخص (میکروسپورا،میکروسپوریدیا).
تفاوت در سازماندهی سلولهای پروکاریوتی و یوکاریوتی در جدول ارائه شده است. 2.1.
جدول 2.1.نشانه های یک سلول پروکاریوتی و یوکاریوتی
2.2. طبقه بندی و مورفولوژی باکتری ها
اصطلاح "باکتری" از این کلمه گرفته شده است باکتری ها،چوب یعنی چی باکتری ها پروکاریوت هستند. آنها به دو حوزه تقسیم می شوند: باکتری هاو آرکی باکتری هاباکتری های موجود در دامنه آرکی باکتری ها،نشان دهنده یکی از قدیمی ترین اشکال زندگی است. آنها ویژگی های ساختاری دیواره سلولی (آنها فاقد پپتیدوگلیکان) و RNA ریبوزومی هستند. هیچ پاتوژن بیماری های عفونی در بین آنها وجود ندارد.
در یک دامنه، باکتری ها به دسته های طبقه بندی زیر تقسیم می شوند: طبقه، شاخه، ردیف، خانواده، جنس، گونه. یکی از دسته بندی های اصلی طبقه بندی می باشد گونه ها.یک گونه مجموعه ای از افراد با منشاء و ژنوتیپ یکسان است که با ویژگی های مشابهی که آنها را از سایر نمایندگان جنس متمایز می کند متحد شده اند. نام گونه مربوط به نامگذاری دوتایی است، یعنی. از دو کلمه تشکیل شده است به عنوان مثال، عامل بیماری دیفتری به صورت نوشته شده است کورینه باکتریوم دیفتری.کلمه اول نام جنس است و با حرف بزرگ نوشته می شود، کلمه دوم نشان دهنده گونه و با حروف کوچک نوشته می شود.
هنگامی که یک گونه دوباره ذکر می شود، نام عمومی به حرف اولیه آن مخفف می شود، به عنوان مثال. C. دیفتری.
مجموعه ای از میکروارگانیسم های همگن جدا شده بر روی یک محیط غذایی که با خواص مورفولوژیکی، رنگی (ارتباط با رنگ ها)، فرهنگی، بیوشیمیایی و آنتی ژنی مشابه مشخص می شود. فرهنگ نابکشت خالص میکروارگانیسم های جدا شده از یک منبع خاص و متفاوت از سایر اعضای گونه نامیده می شود. نژاد.نزدیک به مفهوم "فشار" مفهوم "کلون" است. کلون مجموعه ای از نوادگان است که از یک سلول میکروبی منفرد رشد می کنند.
برای تعیین مجموعه خاصی از میکروارگانیسم ها که از نظر ویژگی های خاص متفاوت هستند، از پسوند "var" (تنوع) استفاده می شود، بنابراین میکروارگانیسم ها بسته به ماهیت تفاوت ها، به عنوان مورفووار (تفاوت در مورفولوژی)، محصولات مقاوم (تفاوت در مقاومت) تعیین می شوند. به عنوان مثال، به آنتی بیوتیک ها)، سرووارها (تفاوت در آنتی ژن ها)، فاگووارها (تفاوت در حساسیت به باکتریوفاژها)، بیووارها (تفاوت در خواص بیولوژیکی)، کیمووارها (تفاوت در خواص بیوشیمیایی) و غیره.
پیش از این، اساس طبقه بندی باکتری ها ویژگی ساختاری دیواره سلولی بود. تقسیم باکتری ها بر اساس ویژگی های ساختاری دیواره سلولی با تغییر رنگ احتمالی آنها در یک رنگ یا دیگری با استفاده از روش گرم همراه است. بر اساس این روش که در سال 1884 توسط دانشمند دانمارکی H. Gram پیشنهاد شد، بسته به نتایج رنگ آمیزی، باکتری ها به گرم مثبت، آبی-بنفش رنگ آمیزی شده و گرم منفی با رنگ قرمز تقسیم می شوند.
در حال حاضر، طبقهبندی بر اساس درجه ارتباط ژنتیکی، بر اساس مطالعه ساختار ژنوم RNA ریبوزومی (rRNA) (به فصل 5 مراجعه کنید)، تعیین درصد جفتهای سیتوزین گوانین (جفتهای GC) در ژنوم، و ساخت یک نقشه محدودیت ژنوم و مطالعه درجه هیبریداسیون. شاخص های فنوتیپی نیز در نظر گرفته می شود: نگرش به رنگ آمیزی گرم، خواص مورفولوژیکی، فرهنگی و بیوشیمیایی، ساختار آنتی ژنی.
دامنه باکتری هاشامل 23 نوع است که موارد زیر از اهمیت پزشکی برخوردار است.
اکثر باکتری های گرم منفی در شاخه گروه بندی می شوند پروتئوباکتری ها(به نام خدای یونانی پروتئوس،قادر به گرفتن اشکال مختلف). تایپ کنید پروتئوباکتری هابه 5 کلاس تقسیم می شود:
کلاس آلفاپروتئوباکتری ها(تولد Rickettsia، Orientia، Erlichia، Bartonella، Brucella)؛
کلاس بتاپروتئوباکتری ها(تولد Bordetella، Burholderia، Neisseria، Spirillum)؛
کلاس گاماپروتئوباکتری ها(نمایندگان خانواده انتروباکتریاسهزایمان فرانسیلا، لژیونلا، کوکسیلا، سودوموناس، ویبریو)؛
کلاس دلتاپروتئوباکتری ها(جنس بیلوفیلا)؛
کلاس اپسیلون پروتئوباکتری(تولد کمپیلوباکتر، هلیکوباکتر).باکتری های گرم منفی نیز شامل انواع زیر هستند:
نوع کلامیدیا(تولد کلامیدیا، کلامیدوفیلا)نوع اسپیروکت ها(تولد Spirocheta، Borrelia، Treponema، Leptospira)؛نوع باکتریوئیدها(تولد باکتریوئیدها، پرووتلا، پورفیروموناس).
باکتری های گرم مثبت در انواع زیر وجود دارند:
تایپ کنید فیرمیکوت هاشامل کلاس است کلستریدیوم(تولد کلستریدیوم، پپتوکوک)کلاس باسیل ها (لیستریا، استافیلوکوک، لاکتوباسیلوس، استرپتوکوک)و کلاس مولیکوت ها(تولد مایکوپلاسما، اورهاپلاسما)که باکتری هایی هستند که دیواره سلولی ندارند.
نوع اکتینوباکتری ها(تولد اکتینومایسس، میکروکوکوس، کورینه باکتریوم، مایکوباکتریوم، گاردنرلا، بیفیدوباکتریوم، پروپیونی باکتریوم، موبیلونکوس).
2.2.1. اشکال مورفولوژیکی باکتری ها
چندین شکل اصلی باکتری وجود دارد: کوکوئید، میله ای شکل، پیچ خورده و منشعب (شکل 2.1).
اشکال کروی یا کوکسی- باکتری های کروی به اندازه 0.5-1 میکرون که با توجه به موقعیت نسبی خود به میکروکوک، دیپلوکوک، استرپتوکوک، تتراکوک، سارسینا و استافیلوکوک تقسیم می شوند.
میکروکوکی (از یونانی. میکرو- کوچک) - سلول های جداگانه قرار دارند.
دیپلوکوکی (از یونانی. دیپلوها- کوکسی های دوتایی) یا جفتی، به صورت جفت (پنوموکوک، گنوکوک، مننگوکوک) قرار دارند، زیرا سلول ها پس از تقسیم جدا نمی شوند. پنوموکوک (عامل بیماری ذات الریه) در طرفین هم شکل نیزه ای دارد و گونوکوک (عامل سوزاک) و مننگوکوک (عامل بیماری سوزاک)
 برنج. 2.1.شکل باکتری ها
برنج. 2.1.شکل باکتری ها
عامل مننژیت اپیدمی) شکل دانه های قهوه با سطح مقعر آنها رو به روی یکدیگر است.
استرپتوکوک (از یونانی. استرپتوز- زنجیره) - سلول هایی به شکل گرد یا کشیده که به دلیل تقسیم سلولی در همان صفحه زنجیره ای را تشکیل می دهند و ارتباط بین آنها را در محل تقسیم حفظ می کنند.
Sarcins (از لات. سارسینا- دسته، عدل) به شکل بسته های 8 کوکسی یا بیشتر چیده شده اند، زیرا آنها در طول تقسیم سلولی در سه صفحه متقابل عمود بر هم تشکیل می شوند.
استافیلوکوک (از یونانی. استافیل- خوشه انگور) - کوکسی که به صورت خوشه انگور در نتیجه تقسیم در صفحات مختلف قرار دارد.
باکتری های میله ای شکلدر اندازه، شکل انتهای سلول و موقعیت نسبی سلول ها متفاوت است. طول سلول 1-10 میکرومتر، ضخامت 0.5-2 میکرومتر است. میله ها می توانند درست باشند
(اشریشیا کلی و غیره) و شکل چماقی نامنظم (کورین باکتریا و غیره). کوچکترین باکتری های میله ای شکل شامل ریکتزیا است.
انتهای میله ها می تواند بریده شود (باسیل سیاه زخم)، گرد (اشریشیا کلی)، نوک تیز (فوزوباکتریا) یا به شکل ضخیم شونده. در مورد دوم، میله مانند یک چماق (Corynebacterium diphtheria) به نظر می رسد.
میله های کمی خمیده ویبریو (Vibrio cholerae) نامیده می شوند. بیشتر باکتری های میله ای شکل به طور تصادفی مرتب می شوند زیرا سلول ها پس از تقسیم از هم جدا می شوند. اگر پس از تقسیم، سلول ها توسط قطعات مشترک دیواره سلولی به هم متصل باقی بمانند و از هم جدا نشوند، در این صورت در زاویه ای نسبت به یکدیگر قرار می گیرند (کورین باکتریوم دیفتری) یا تشکیل یک زنجیره (باسیل سیاه زخم) می دهند.
شکل های پیچ خورده- باکتری های مارپیچی شکل که در دو نوع هستند: اسپیرلا و اسپیروکت. اسپیرلاها ظاهر سلول های پیچ خورده چوب پنبه ای شکل با فرهای بزرگ دارند. اسپیریل های بیماری زا شامل عامل ایجاد کننده سودوکو (بیماری نیش موش) و همچنین کمپیلوباکتر و هلیکوباکتر هستند که انحناهایی شبیه بال های مرغ دریایی در حال پرواز دارند. اسپیروکتها باکتریهای نازک، دراز و پیچخوردهای هستند که در پیچهای کوچکتر و الگوی حرکت با اسپیرلا متفاوت هستند. ویژگی های ساختار آنها در زیر توضیح داده شده است.
انشعاب -باکتری های میله ای شکل، که می توانند شاخه های Y شکل در بیفیدوباکتری ها داشته باشند، همچنین می توانند به شکل سلول های شاخه دار رشته ای ارائه شوند که می توانند در هم تنیده شوند و میسلیوم را تشکیل دهند، همانطور که در اکتینومیست ها مشاهده می شود.
2.2.2. ساختار سلولی باکتریایی
ساختار باکتری ها با استفاده از میکروسکوپ الکترونی سلول های کامل و بخش های فوق نازک آنها و همچنین روش های دیگر به خوبی مورد مطالعه قرار گرفته است. سلول باکتری توسط یک غشاء متشکل از یک دیواره سلولی و یک غشای سیتوپلاسمی احاطه شده است. در زیر پوسته پروتوپلاسم وجود دارد، متشکل از سیتوپلاسم با اجزاء و یک دستگاه ارثی - آنالوگ هسته، به نام نوکلوئید (شکل 2.2). ساختارهای اضافی وجود دارد: کپسول، میکروکپسول، مخاط، تاژک، پیلی. برخی از باکتری ها در شرایط نامساعد قادر به تشکیل هاگ هستند.
 برنج. 2.2.ساختار یک سلول باکتریایی: 1 - کپسول. 2 - دیواره سلولی 3 - غشای سیتوپلاسمی; 4 - مزوزوم ها; 5 - نوکلوئید؛ 6 - پلاسمید; 7 - ریبوزوم; 8 - شمول; 9 - تاژک؛ 10 - پیلی (پرز)
برنج. 2.2.ساختار یک سلول باکتریایی: 1 - کپسول. 2 - دیواره سلولی 3 - غشای سیتوپلاسمی; 4 - مزوزوم ها; 5 - نوکلوئید؛ 6 - پلاسمید; 7 - ریبوزوم; 8 - شمول; 9 - تاژک؛ 10 - پیلی (پرز)
دیواره سلولی- ساختاری قوی و الاستیک که به باکتری شکل خاصی می دهد و همراه با غشای سیتوپلاسمی زیرین، فشار اسمزی بالا را در سلول باکتری مهار می کند. در فرآیند تقسیم سلولی و انتقال متابولیت ها نقش دارد، دارای گیرنده هایی برای باکتریوفاژها، باکتریوسین ها و مواد مختلف است. ضخیم ترین دیواره سلولی در باکتری های گرم مثبت یافت می شود (شکل 2.3). بنابراین، اگر ضخامت دیواره سلولی باکتری های گرم منفی حدود 15-20 نانومتر باشد، در باکتری های گرم مثبت می تواند به 50 نانومتر یا بیشتر برسد.
اساس دیواره سلولی باکتری است پپتیدوگلیکانپپتیدوگلیکان یک پلیمر است. آن را با زنجیره های گلیکان پلی ساکارید موازی متشکل از تکرار N-acetylglucosamine و N-acetylmuramic اسید که توسط یک پیوند گلیکوزیدی متصل شده اند، نشان داده می شود. این پیوند توسط لیزوزیم که یک استیل مورامیداز است شکسته می شود.
یک تتراپپتید توسط پیوندهای کووالانسی به N-استیل مورامیک اسید متصل می شود. تتراپپتید متشکل از ال-آلانین است که به N-استیل مورامیک اسید متصل است. دی گلوتامین که در باکتری های گرم مثبت با ال لیزین ترکیب می شود و در گرم تری
 برنج. 2.3.طرح معماری دیواره سلولی باکتری
برنج. 2.3.طرح معماری دیواره سلولی باکتری
باکتری های مفید - با اسید دی آمینوپیملیک (DAP)، که پیش ساز لیزین در فرآیند بیوسنتز باکتریایی اسیدهای آمینه است و یک ترکیب منحصر به فرد است که فقط در باکتری ها وجود دارد. اسید آمینه چهارم D-آلانین است (شکل 2.4).
دیواره سلولی باکتری های گرم مثبت حاوی مقادیر کمی پلی ساکارید، لیپید و پروتئین است. جزء اصلی دیواره سلولی این باکتری ها پپتیدوگلیکان چندلایه (مورئین، موکوپپتید) است که 90-40 درصد جرم دیواره سلولی را تشکیل می دهد. تتراپپتیدهای لایه های مختلف پپتیدوگلیکان در باکتری های گرم مثبت توسط زنجیره های پلی پپتیدی از 5 باقیمانده گلیسین (پنتاگلیسین) به یکدیگر متصل می شوند که به پپتیدوگلیکان ساختار هندسی سفت و سختی می دهد (شکل 2.4، b). پیوند کووالانسی با پپتیدوگلیکان دیواره سلولی باکتری های گرم مثبت اسیدهای تیکوئیک(از یونانی تکوس- دیوار) که مولکول های آن زنجیره های 50-8 گلیسرول و باقی مانده های ریبیتول است که توسط پل های فسفات به هم متصل شده اند. شکل و قدرت باکتری ها توسط ساختار فیبری سفت و سخت پپتیدوگلیکان چند لایه، با پیوندهای متقابل پپتیدها داده می شود.
 برنج. 2.4.ساختار پپتیدوگلیکان: a - باکتری های گرم منفی. ب - باکتری های گرم مثبت
برنج. 2.4.ساختار پپتیدوگلیکان: a - باکتری های گرم منفی. ب - باکتری های گرم مثبت
توانایی باکتری های گرم مثبت در حفظ بنفشه جنتین در ترکیب با ید هنگام رنگ آمیزی با استفاده از رنگ آمیزی گرم (رنگ آبی-بنفش باکتری ها) با خاصیت پپتیدوگلیکان چند لایه برای تعامل با رنگ مرتبط است. علاوه بر این، درمان بعدی اسمیر باکتریایی با الکل باعث باریک شدن منافذ در پپتیدوگلیکان و در نتیجه حفظ رنگ در دیواره سلولی می شود.
باکتری های گرم منفی پس از قرار گرفتن در معرض الکل، رنگ را از دست می دهند، که به دلیل مقدار کمتر پپتیدوگلیکان (5-10٪ از توده دیواره سلولی) است. آنها با الکل تغییر رنگ داده و هنگامی که با فوشین یا سافرانین درمان می شوند قرمز می شوند. این به دلیل ویژگی های ساختاری دیواره سلولی است. پپتیدوگلیکان در دیواره سلولی باکتری های گرم منفی با 1-2 لایه نشان داده می شود. تتراپپتیدهای لایه ها توسط یک پیوند پپتیدی مستقیم بین گروه آمینه DAP یک تتراپپتید و گروه کربوکسیل D-آلانین تتراپپتید لایه دیگر به یکدیگر متصل می شوند (شکل 2.4، a). در خارج از پپتیدوگلیکان یک لایه وجود دارد لیپوپروتئین،از طریق DAP به پپتیدوگلیکان متصل می شود. به دنبال غشای خارجیدیواره سلولی
غشای خارجییک ساختار موزاییکی است که از لیپوپلی ساکاریدها (LPS)، فسفولیپیدها و پروتئین ها تشکیل شده است. لایه داخلی آن با فسفولیپیدها نشان داده می شود و لایه بیرونی حاوی LPS است (شکل 2.5). بنابراین، مم بیرونی
 برنج. 2.5.ساختار لیپوپلی ساکارید
برنج. 2.5.ساختار لیپوپلی ساکارید
بران نامتقارن است. غشای خارجی LPS از سه قطعه تشکیل شده است:
لیپید A ساختار محافظه کارانه ای دارد که در باکتری های گرم منفی تقریباً یکسان است. لیپید A شامل واحدهای دی ساکارید گلوکزآمین فسفریله شده است که زنجیره های بلند اسیدهای چرب به آنها متصل است (شکل 2.5 را ببینید).
هسته، یا هسته، بخش پوسته (از lat. هسته- هسته)، ساختار الیگوساکارید نسبتا محافظه کار.
یک زنجیره پلی ساکارید اختصاصی O بسیار متغیر که با تکرار توالی های اولیگوساکاریدی یکسان تشکیل شده است.
LPS توسط لیپید A در غشای خارجی لنگر انداخته است که باعث سمیت LPS می شود و بنابراین با اندوتوکسین شناسایی می شود. تخریب باکتری ها توسط آنتی بیوتیک ها منجر به ترشح مقادیر زیادی اندوتوکسین می شود که می تواند باعث ایجاد شوک اندوتوکسیک در بیمار شود. هسته یا بخش اصلی LPS از لیپید A گسترش می یابد. ثابت ترین قسمت هسته LPS کتودوکسیوکتونیک اسید است. زنجیره پلی ساکارید اختصاصی O که از هسته مولکول LPS گسترش می یابد،
متشکل از واحدهای تکراری الیگوساکارید، سروگروپ، سرووار (نوعی باکتری که با استفاده از سرم ایمنی تشخیص داده می شود) سویه خاصی از باکتری را تعیین می کند. بنابراین، مفهوم LPS با مفهوم O-آنتی ژن همراه است که توسط آن باکتری ها را می توان متمایز کرد. تغییرات ژنتیکی می تواند منجر به نقص، کوتاه شدن LPS باکتریایی و در نتیجه ظهور کلونی های خشن از فرم های R شود که ویژگی آنتی ژن O را از دست می دهند.
همه باکتری های گرم منفی دارای یک زنجیره پلی ساکارید O-specific کامل نیستند که از واحدهای الیگوساکاریدی تکرار شونده تشکیل شده است. به ویژه، باکتری های جنس نایسریایک گلیکولیپید کوتاه به نام لیپولیگوساکارید (LOS) دارند. قابل مقایسه با فرم R است که ویژگی آنتی ژن O را از دست داده است، مشاهده شده در سویه های خشن جهش یافته E. coli.ساختار VOC شبیه ساختار گلیکوسفنگولیپید غشای سیتوپلاسمی انسان است، بنابراین VOC از میکروب تقلید می کند و به آن اجازه می دهد از پاسخ ایمنی میزبان فرار کند.
پروتئین های ماتریکس غشای خارجی به گونه ای در آن نفوذ می کنند که مولکول های پروتئینی به نام پورینامی،حفره های آبدوست مرزی که آب و مولکول های کوچک آبدوست با جرم نسبی تا 700 دسیونیت از آن عبور می کنند.
بین غشای خارجی و سیتوپلاسمی قرار دارد فضای پری پلاسمیک،یا آنزیم های حاوی پری پلاسم (پروتئازها، لیپازها، فسفاتازها، نوکلئازها، بتالاکتامازها) و همچنین اجزای سیستم های انتقال.
هنگامی که سنتز دیواره سلولی باکتری تحت تأثیر لیزوزیم، پنی سیلین، عوامل محافظ بدن و سایر ترکیبات مختل می شود، سلول هایی با شکل تغییر یافته (اغلب کروی) تشکیل می شوند: پروتوپلاست ها- باکتری هایی که به طور کامل فاقد دیواره سلولی هستند. کروپلاست ها- باکتری با دیواره سلولی تا حدی حفظ شده. پس از حذف بازدارنده دیواره سلولی، چنین باکتری های تغییر یافته می توانند معکوس شوند، به عنوان مثال. یک دیواره سلولی کامل به دست آورد و شکل اولیه خود را بازیابی کرد.
باکتری هایی از نوع کروی یا پروتوپلاست که تحت تأثیر آنتی بیوتیک ها یا عوامل دیگر توانایی سنتز پپتیدوگلیکان را از دست داده و قادر به تولید مثل هستند، نامیده می شوند. L شکل ها(از نام مؤسسه D. Lister، جایی که آنها ابتدا
مطالعه شده اند). شکل L همچنین می تواند در نتیجه جهش ایجاد شود. آنها سلولهای کروی و فلاسکی شکل با اندازه های مختلف از نظر اسمزی حساس هستند، از جمله آنهایی که از فیلترهای باکتریایی عبور می کنند. برخی از اشکال L (ناپایدار)، زمانی که عاملی که منجر به تغییرات در باکتریها میشود حذف شود، میتوانند معکوس شوند و به سلول اصلی باکتری برگردند. اشکال L می توانند توسط بسیاری از پاتوژن های بیماری های عفونی تولید شوند.
غشای سیتوپلاسمیدر میکروسکوپ الکترونی مقاطع فوق نازک، یک غشای سه لایه است (2 لایه تیره، هر کدام 2.5 نانومتر ضخامت، که توسط یک لایه میانی سبک از هم جدا شده اند). از نظر ساختار، شبیه پلاسمالمای سلول های حیوانی است و از یک لایه دوگانه از لیپیدها، عمدتاً فسفولیپیدها، با سطح جاسازی شده و پروتئین های انتگرال تشکیل شده است که به نظر می رسد از طریق ساختار غشاء نفوذ می کنند. برخی از آنها پرمازهایی هستند که در حمل و نقل مواد نقش دارند. برخلاف سلول های یوکاریوتی، غشای سیتوپلاسمی سلول باکتری فاقد استرول است (به استثنای مایکوپلاسما).
غشای سیتوپلاسمی یک ساختار پویا با اجزای متحرک است، بنابراین به عنوان یک ساختار سیال متحرک در نظر گرفته می شود. این قسمت بیرونی سیتوپلاسم باکتری ها را احاطه کرده و در تنظیم فشار اسمزی، انتقال مواد و متابولیسم انرژی سلول (به دلیل آنزیم های زنجیره انتقال الکترون، آدنوزین تری فسفاتاز - ATPase و غیره) نقش دارد. با رشد بیش از حد (در مقایسه با رشد دیواره سلولی)، غشای سیتوپلاسمی شکل می گیرد invaginates - invaginations به شکل ساختارهای غشایی پیچیده پیچیده، به نام مزوزوم هاساختارهای پیچ خورده کمتر غشاهای داخل سیتوپلاسمی نامیده می شوند. نقش مزوزوم ها و غشاهای داخل سیتوپلاسمی به طور کامل شناخته نشده است. حتی پیشنهاد می شود که آنها مصنوع هستند که پس از آماده سازی (تثبیت) یک نمونه برای میکروسکوپ الکترونی رخ می دهد. با این وجود، اعتقاد بر این است که مشتقات غشای سیتوپلاسمی در تقسیم سلولی شرکت می کنند و انرژی برای سنتز دیواره سلولی فراهم می کنند و در ترشح مواد، اسپورزایی، یعنی شرکت می کنند. در فرآیندهایی با مصرف انرژی بالا سیتوپلاسم حجم اصلی باکتری ها را اشغال می کند
سلول و متشکل از پروتئین های محلول، اسیدهای ریبونوکلئیک، اجزاء و گرانول های کوچک متعدد - ریبوزوم ها، مسئول سنتز (ترجمه) پروتئین ها است.
ریبوزوم هابر خلاف ریبوزوم های 80S مشخصه سلول های یوکاریوتی، اندازه باکتری ها حدود 20 نانومتر و ضریب رسوب 70S است. بنابراین، برخی از آنتی بیوتیک ها با اتصال به ریبوزوم های باکتری، سنتز پروتئین باکتریایی را بدون تأثیر بر سنتز پروتئین در سلول های یوکاریوتی مهار می کنند. ریبوزوم های باکتریایی می توانند به دو زیر واحد تقسیم شوند: 50S و 30S. rRNA یک عنصر حفاظت شده از باکتری ها ("ساعت مولکولی" تکامل) است. 16S rRNA بخشی از زیر واحد ریبوزومی کوچک و 23S rRNA بخشی از زیر واحد ریبوزومی بزرگ است. مطالعه 16S rRNA اساس سیستماتیک ژن است که به فرد اجازه می دهد تا درجه ارتباط ارگانیسم ها را ارزیابی کند.
سیتوپلاسم حاوی اجزاء مختلفی به شکل گرانول های گلیکوژن، پلی ساکاریدها، اسید β-هیدروکسی بوتیریک و پلی فسفات ها (ولوتین) است. آنها زمانی که مواد مغذی زیادی در محیط وجود دارد تجمع می یابند و به عنوان مواد ذخیره ای برای نیازهای تغذیه و انرژی عمل می کنند.
وولیوتینمیل ترکیبی به رنگ های اساسی دارد و با استفاده از روش های رنگ آمیزی خاص (مثلاً طبق گفته نایسر) به راحتی به صورت گرانول های متاکروماتیک تشخیص داده می شود. با تولویدین بلو یا متیلن بلو، ولوتین به رنگ قرمز-بنفش رنگ آمیزی می شود و سیتوپلاسم باکتری به رنگ آبی رنگ می شود. آرایش مشخصه گرانول های ولوتین در باسیل دیفتری به شکل قطب های سلولی به شدت رنگ آمیزی شده آشکار می شود. رنگ متاکروماتیک ولوتین با محتوای بالای پلی فسفات معدنی پلیمریزه شده مرتبط است. در زیر میکروسکوپ الکترونی، آنها مانند گرانول های الکترونی متراکم به اندازه 0.1-1 میکرون به نظر می رسند.
نوکلوئید- معادل هسته در باکتری ها. این در ناحیه مرکزی باکتری ها به شکل DNA دو رشته ای قرار دارد که به طور محکم مانند یک توپ بسته بندی شده است. نوکلوئید باکتری ها بر خلاف یوکاریوت ها دارای پوشش هسته ای، هسته و پروتئین های اساسی (هیستون ها) نیستند. اکثر باکتری ها حاوی یک کروموزوم هستند که توسط یک مولکول DNA بسته شده در یک حلقه نشان داده می شود. اما برخی از باکتری ها دارای دو کروموزوم حلقه ای شکل هستند (V. cholerae)و کروموزوم های خطی (به بخش 5.1.1 مراجعه کنید). این نوکلوئید پس از رنگ آمیزی با لکه های اختصاصی DNA در میکروسکوپ نوری آشکار می شود
روش ها: به گفته Feulgen یا به گفته Romanovsky-Gimsa. در الگوهای پراش الکترونی بخشهای بسیار نازک باکتریها، نوکلوئید بهعنوان مناطق نورانی با ساختارهای رشتهای و نخ مانند DNA که در نواحی خاصی به غشای سیتوپلاسمی یا مزوزوم درگیر در همانندسازی کروموزوم متصل شدهاند، ظاهر میشود.
علاوه بر نوکلوئید، سلول باکتری حاوی فاکتورهای وراثت خارج کروموزومی است - پلاسمیدها (به بخش 5.1.2 مراجعه کنید)، که حلقه های بسته کووالانسی DNA هستند.
کپسول، میکروکپسول، موکوس.کپسول -ساختار مخاطی با ضخامت بیش از 0.2 میکرون، به طور محکم با دیواره سلولی باکتری مرتبط است و دارای مرزهای خارجی مشخص است. کپسول در اسمیرهای اثری از مواد پاتولوژیک قابل مشاهده است. در کشت های خالص باکتریایی، کپسول کمتر تشکیل می شود. با استفاده از روش های خاص رنگ آمیزی اسمیر مطابق Burri-Gins تشخیص داده می شود که کنتراست منفی مواد کپسول ایجاد می کند: جوهر زمینه تیره ای را در اطراف کپسول ایجاد می کند. کپسول از پلی ساکاریدها (اگزوپلی ساکاریدها) و گاهی اوقات از پلی پپتیدها تشکیل شده است، به عنوان مثال، در باسیل سیاه زخم از پلیمرهای D-گلوتامیک اسید تشکیل شده است. کپسول آبدوست و حاوی مقدار زیادی آب است. از فاگوسیتوز باکتری ها جلوگیری می کند. کپسول آنتی ژنی است: آنتی بادی های کپسول باعث بزرگ شدن آن می شود (واکنش تورم کپسول).
بسیاری از باکتری ها تشکیل می شوند میکروکپسول- تشکیل مخاطی با ضخامت کمتر از 0.2 میکرون که فقط با میکروسکوپ الکترونی قابل تشخیص است.
باید از کپسول متمایز شود لجن -اگزوپلی ساکاریدهای مخاطی که مرزهای خارجی مشخصی ندارند. مخاط در آب محلول است.
اگزوپلی ساکاریدهای موکوئیدی مشخصه سویه های موکوییدی سودوموناس آئروژینوزا هستند که اغلب در خلط بیماران مبتلا به فیبروز کیستیک یافت می شوند. اگزوپلی ساکاریدهای باکتریایی در چسبندگی (چسبیدن به بسترها) نقش دارند. آنها گلیکوکالیکس نیز نامیده می شوند.
کپسول و مخاط از باکتری ها در برابر آسیب و خشک شدن محافظت می کند، زیرا آب دوست بودن، آب را به خوبی می چسباند و از عملکرد عوامل محافظتی ماکرو ارگانیسم ها و باکتریوفاژها جلوگیری می کند.
تاژکباکتری ها تحرک سلول باکتری را تعیین می کنند. تاژک ها رشته های نازکی هستند که به خود می گیرند
آنها از غشای سیتوپلاسمی سرچشمه می گیرند و از خود سلول بلندتر هستند. ضخامت تاژک 12-20 نانومتر، طول 3-15 میکرومتر است. آنها از سه بخش تشکیل شده اند: یک رشته مارپیچ، یک قلاب و یک بدنه پایه حاوی میله ای با دیسک های خاص (یک جفت دیسک در باکتری های گرم مثبت و دو جفت در باکتری های گرم منفی). تاژک ها توسط دیسک ها به غشای سیتوپلاسمی و دیواره سلولی متصل می شوند. این باعث ایجاد اثر یک موتور الکتریکی با یک میله - یک روتور - می شود که تاژک را می چرخاند. اختلاف پتانسیل پروتون در غشای سیتوپلاسمی به عنوان منبع انرژی استفاده می شود. مکانیسم چرخش توسط پروتون سنتتاز ATP ارائه می شود. سرعت چرخش تاژک می تواند به 100 دور در ثانیه برسد. اگر یک باکتری چندین تاژک داشته باشد، آنها شروع به چرخش همزمان می کنند، در یک بسته نرم افزاری در هم تنیده می شوند و نوعی پروانه را تشکیل می دهند.
تاژک ها از پروتئینی به نام فلاژلین ساخته شده اند. (فلاژلوم- تاژک)، که یک آنتی ژن است - به اصطلاح آنتی ژن H. زیر واحدهای فلاژلین به صورت مارپیچی پیچ خورده اند.
تعداد تاژک ها در گونه های مختلف باکتری از یک (monotrichus) در Vibrio cholerae تا ده ها و صدها تاژک در امتداد محیط باکتری (peritrichus)، در Escherichia coli، Proteus و غیره متغیر است. از سلول آمفیتریشی دارای یک تاژک یا دسته ای از تاژک در انتهای مخالف سلول است.
تاژک ها با استفاده از میکروسکوپ الکترونی فراورده های پوشش داده شده با فلزات سنگین، یا در میکروسکوپ نوری پس از درمان با روش های خاص مبتنی بر اچ و جذب مواد مختلف که منجر به افزایش ضخامت تاژک می شود (به عنوان مثال، پس از نقره سازی) شناسایی می شوند.
پرز، یا پیلی (فیمبریا)- تشکیلات نخ مانند، نازک تر و کوتاه تر (3-10 نانومتر * 0.3-10 میکرومتر) از تاژک. پیلی ها از سطح سلول گسترش یافته و از پروتئین پیلین تشکیل شده اند. انواع مختلفی از پیلی شناخته شده است. نوع عمومی پیلی مسئول اتصال به بستر، تغذیه و متابولیسم آب و نمک است. آنها متعدد هستند - چند صد در هر سلول. جنس پیلی (1-3 در هر سلول) تماس بین سلول ها را ایجاد می کند و اطلاعات ژنتیکی را بین آنها با صرف انتقال می دهد (به فصل 5 مراجعه کنید). جالب توجه خاص پیلی نوع IV است که انتهای آن آبگریز است و در نتیجه پیچ می خورد؛ به این پیلی ها فر نیز می گویند. محل
آنها در قطب های سلول قرار دارند. این پیلی ها در باکتری های بیماری زا یافت می شوند. آنها خواص آنتی ژنی دارند، باکتری ها را با سلول میزبان در تماس می گیرند و در تشکیل بیوفیلم شرکت می کنند (به فصل 3 مراجعه کنید). بسیاری از پیلی ها گیرنده های باکتریوفاژها هستند.
اختلافات -شکل عجیبی از باکتری های در حال استراحت با نوع گرم مثبت ساختار دیواره سلولی. باکتری های اسپورساز این جنس باسیل،که در آن اندازه هاگ از قطر سلول بیشتر نباشد باسیل نامیده می شود. باکتری های اسپورساز که در آنها اندازه هاگ از قطر سلول بیشتر است و به همین دلیل شکل دوک به خود می گیرند، نامیده می شوند. کلستریدیا،به عنوان مثال باکتری های جنس کلستریدیوم(از لات کلستریدیوم- دوک). هاگ ها در برابر اسید مقاوم هستند، بنابراین با استفاده از روش Aujeszky یا Ziehl-Neelsen قرمز رنگ می شوند و سلول رویشی به رنگ آبی رنگ می شود.
هاگ زایی، شکل و محل قرارگیری هاگ ها در یک سلول (روشی) از ویژگی های گونه ای باکتری ها است که به آنها اجازه می دهد تا از یکدیگر متمایز شوند. شکل هاگ ها می تواند بیضی یا کروی باشد، مکان در سلول انتهایی است، یعنی. در انتهای چوب (در عامل ایجاد کننده کزاز)، زیر ترمینال - نزدیک به انتهای چوب (در عوامل ایجاد کننده بوتولیسم، گانگرن گازی) و مرکزی (در باسیل سیاه زخم).
فرآیند اسپورزایی (اسپورزایی) از چندین مرحله می گذرد که طی آن بخشی از سیتوپلاسم و کروموزوم سلول رویشی باکتری از هم جدا می شود و توسط یک غشای سیتوپلاسمی در حال رشد احاطه شده است - یک پروسپور تشکیل می شود.
پروتوپلاست پروسپور حاوی یک نوکلوئید، یک سیستم سنتز پروتئین و یک سیستم تولید انرژی مبتنی بر گلیکولیز است. سیتوکروم ها حتی در هوازی نیز وجود ندارند. حاوی ATP نیست، انرژی برای جوانه زنی به شکل 3 گلیسرول فسفات ذخیره می شود.
پروسپور توسط دو غشای سیتوپلاسمی احاطه شده است. لایه احاطه کننده غشای داخلی اسپور نامیده می شود دیواره هاگ،از پپتیدوگلیکان تشکیل شده و منبع اصلی دیواره سلولی در طول جوانه زنی هاگ است.
بین غشای خارجی و دیواره هاگ، یک لایه ضخیم متشکل از پپتیدوگلیکان تشکیل می شود که دارای پیوندهای متقابل زیادی است - قشر
خارج از غشای سیتوپلاسمی خارجی قرار دارد پوسته هاگ،متشکل از پروتئین های کراتین مانند،
نگه داشتن پیوندهای دی سولفیدی درون مولکولی متعدد این پوسته در برابر عوامل شیمیایی مقاومت می کند. هاگ های برخی از باکتری ها دارای پوشش اضافی هستند - اگزوسپوریومطبیعت لیپوپروتئینی به این ترتیب، یک پوسته چند لایه با نفوذپذیری ضعیف تشکیل می شود.
هاگ زایی با مصرف شدید پروسپور و سپس پوسته هاگ در حال توسعه اسید دی پی کولینیک و یون های کلسیم همراه است. اسپور مقاومت گرمایی را به دست می آورد که با وجود دی پی کولینات کلسیم در آن همراه است.
اسپور به دلیل وجود پوسته چند لایه، دی پی کولینات کلسیم، محتوای کم آب و فرآیندهای متابولیک کند می تواند برای مدت طولانی باقی بماند. به عنوان مثال، در خاک، پاتوژن های سیاه زخم و کزاز می توانند برای چندین دهه باقی بمانند.
در شرایط مساعد، هاگ ها جوانه می زنند و سه مرحله متوالی را طی می کنند: فعال شدن، شروع، رشد. در این حالت یک باکتری از یک هاگ تشکیل می شود. فعال سازی آمادگی برای جوانه زنی است. در دمای 60-80 درجه سانتیگراد، هاگ برای جوانه زنی فعال می شود. شروع جوانه زنی چند دقیقه طول می کشد. مرحله رشد با رشد سریع همراه با تخریب پوسته و ظهور یک نهال مشخص می شود.
2.2.3. ویژگی های ساختاری اسپیروکت ها، ریکتزیا، کلامیدیا، اکتینومیست ها و مایکوپلاسماها
اسپیروکت ها- باکتری های پیچیده دراز نازک آنها از یک دیواره سلولی غشایی بیرونی تشکیل شده اند که یک استوانه سیتوپلاسمی را احاطه کرده است. در بالای غشای خارجی یک پوشش شفاف از طبیعت گلیکوزامینوگلیکان وجود دارد. در زیر غشای خارجی دیواره سلولی فیبرهایی وجود دارد که به دور استوانه سیتوپلاسمی می پیچند و به باکتری ها شکل مارپیچ می دهند. فیبریل ها به انتهای سلول متصل شده و به سمت یکدیگر هدایت می شوند. تعداد و آرایش فیبریل ها در بین گونه ها متفاوت است. فیبریل ها در حرکت اسپیروکت ها نقش دارند و به سلول ها حرکت چرخشی، خمشی و انتقالی می دهند. در این حالت، اسپیروکت ها حلقه ها، فرها و خم هایی را تشکیل می دهند که به آنها فرهای ثانویه می گویند. اسپیروکت ها رنگ ها را به خوبی درک نمی کنند. آنها معمولاً مطابق رومانوفسکی-گیمسا یا روکش نقره نقاشی می شوند. زنده
شکل اسپیروکت با استفاده از میکروسکوپ فاز-کنتراست یا میدان تاریک بررسی می شود.
اسپیروکت ها توسط سه جنس که برای انسان بیماری زا هستند نشان داده می شوند: ترپونما، بورلیا، لپتوسپیرا.
ترپونما(جنس ترپونما)ظاهری نخ های نازک و پیچ خورده با چوب پنبه با 8 تا 12 فر کوچک یکنواخت دارند. در اطراف پروتوپلاست ترپونما 3-4 فیبریل (فلاژلا) وجود دارد. سیتوپلاسم حاوی رشته های سیتوپلاسمی است. نمایندگان بیماری زا هستند T. pallidum- عامل ایجاد کننده سیفلیس، T. pertenue- عامل بیماری استوایی یاوز. ساپروفیت ها نیز وجود دارند - ساکنان حفره دهان انسان و گل و لای مخازن.
بورلیا(جنس بورلیا)،بر خلاف ترپونما، آنها بلندتر هستند، 3-8 فر بزرگ و 7-20 فیبریل دارند. اینها شامل عامل ایجاد کننده تب عود کننده است (B. recurrentis)و پاتوژن های بیماری لایم (V. بورگدورفری)و بیماری های دیگر
لپتوسپیرا(جنس لپتوسپیرا)آنها دارای فرهای کم عمق و مکرر به شکل یک طناب پیچ خورده هستند. انتهای این اسپیروکت ها مانند قلاب هایی خمیده هستند که در انتهای آن ها ضخیم می شوند. با تشکیل فرهای ثانویه، آنها شکل حروف S یا C را به خود می گیرند. دارای دو فیبریل محوری نماینده بیماری زا L. استیضاحهنگامی که از طریق آب یا غذا خورده شود باعث لپتوسپیروز می شود که منجر به خونریزی و یرقان می شود.
ریکتزیا متابولیسمی مستقل از سلول میزبان دارند، با این حال، ممکن است ترکیبات پرانرژی را از سلول میزبان برای تولید مثل خود دریافت کنند. در اسمیرها و بافت ها طبق رومانوفسکی-گیمسا، طبق گفته ماکیاولو-زدودوفسکی (ریکتسیا قرمز و سلول های آلوده آبی هستند) رنگ آمیزی می شوند.
در انسان، ریکتزیا باعث تیفوس همه گیر می شود. (R. prowazekii)،ریکتسیوز ناشی از کنه (R. sibirica)،تب منقوط کوههاى راکى (R. rickettsii)و سایر ریکتزیوزها
ساختار دیواره سلولی آنها شبیه باکتری های گرم منفی است، اگرچه تفاوت هایی وجود دارد. حاوی پپتیدوگلیکان معمولی نیست: کاملاً فاقد N-استیل مورامیک اسید است. دیواره سلولی از یک غشای خارجی دوتایی تشکیل شده است که شامل لیپوپلی ساکارید و پروتئین است. علیرغم عدم وجود پپتیدوگلیکان، دیواره سلولی کلامیدیا سفت و سخت است. سیتوپلاسم سلول توسط غشای سیتوپلاسمی داخلی محدود می شود.
روش اصلی برای تشخیص کلامیدیا رنگ آمیزی رومانوفسکی-گیمسا است. رنگ بستگی به مرحله چرخه زندگی دارد: اجسام ابتدایی در پس زمینه سیتوپلاسم آبی سلول بنفش به نظر می رسند، اجسام مشبک آبی به نظر می رسند.
کلامیدیا در انسان باعث آسیب به چشم ها (تراخم، ورم ملتحمه)، دستگاه ادراری تناسلی، ریه ها و غیره می شود.
اکتینومیست ها- باکتری های گرم مثبت شاخه دار، رشته ای یا میله ای شکل. نام آن (از یونانی. عمل می کند- اشعه، مایکز- قارچ) به دلیل تشکیل دروسن در بافت های آسیب دیده - گرانول های نخ های محکم در هم تنیده به شکل
پرتوهایی که از مرکز گسترش یافته و به ضخامت های فلاسکی شکل ختم می شوند. اکتینومیست ها مانند قارچ ها میسلیوم - سلول های در هم تنیده نخ مانند (هیف) را تشکیل می دهند. آنها میسلیوم سوبسترا را تشکیل می دهند که در نتیجه رشد سلولی در محیط غذایی تشکیل می شود و میسلیوم هوایی که در سطح محیط رشد می کند. اکتینومیست ها می توانند با تکه تکه شدن میسلیوم به سلول هایی شبیه باکتری های میله ای شکل و کوکوئید تقسیم شوند. بر روی هیف های هوایی اکتینومیست ها، هاگ هایی تشکیل می شوند که برای تولید مثل استفاده می شوند. اسپورهای اکتینومیست معمولاً در برابر حرارت مقاوم نیستند.
یک شاخه فیلوژنتیک مشترک با اکتینومیست ها توسط به اصطلاح اکتینومیست های نوکاردیفرم (nocardioform) تشکیل می شود - گروهی از باکتری های میله ای شکل با شکل نامنظم. نمایندگان فردی آنها فرم های شاخه ای را تشکیل می دهند. اینها شامل باکتری های این جنس است کورینه باکتریوم، مایکوباکتریوم، نوکاردیااکتینومیست های شبه نوکاردی با وجود قندهای آرابینوز، گالاکتوز و همچنین اسیدهای مایکولیک و مقادیر زیادی اسیدهای چرب در دیواره سلولی متمایز می شوند. اسیدهای مایکولیک و لیپیدهای دیواره سلولی مقاومت اسیدی باکتری ها، به ویژه مایکوباکتریوم توبرکلوزیس و جذام را تعیین می کنند (در صورت رنگ آمیزی طبق Ziehl-Neelsen، آنها قرمز هستند و باکتری ها و عناصر بافتی غیر مقاوم به اسید، خلط آبی هستند).
اکتینومیست های بیماری زا باعث اکتینومایکوز، نوکاردیا - نوکاردیوز، مایکوباکتریوم - سل و جذام، کورینه باکتریا - دیفتری می شوند. اشکال ساپروفیت اکتینومیست ها و اکتینومیست های نوکاردیفرم در خاک گسترده هستند، بسیاری از آنها تولید کننده آنتی بیوتیک هستند.
مایکوپلاسماها- باکتری های کوچک (0.15-1 میکرومتر) که فقط توسط یک غشای سیتوپلاسمی حاوی استرول احاطه شده اند. آنها متعلق به کلاس هستند مولیکوت هابه دلیل عدم وجود دیواره سلولی، مایکوپلاسماها از نظر اسمزی حساس هستند. آنها اشکال مختلفی دارند: کوکوئید، رشته ای، فلاسکی شکل. این اشکال در میکروسکوپ فاز کنتراست کشت خالص مایکوپلاسما قابل مشاهده است. روی یک محیط مغذی متراکم، مایکوپلاسماها کلونی هایی را تشکیل می دهند که شبیه تخم مرغ های سرخ شده هستند: یک قسمت مات مرکزی غوطه ور در محیط و یک محیط نیمه شفاف به شکل یک دایره.
مایکوپلاسما باعث پنومونی آتیپیک در انسان می شود (مایکوپلاسما پنومونیه)و ضایعات دستگاه تناسلی ادراری
(م. انسانو غیره.). مایکوپلاسما نه تنها در حیوانات، بلکه در گیاهان نیز باعث بیماری می شود. نمایندگان غیر بیماری زا نیز بسیار گسترده هستند.
2.3. ساختار و طبقه بندی قارچ
قارچ متعلق به دامنه است یوکاریا،پادشاهی قارچ (Mycota، Mycetes).اخیراً قارچ ها و تک یاخته ها به پادشاهی های جداگانه تقسیم شده اند: پادشاهی یومیکوتا(قارچ واقعی)، پادشاهی کرومیستاو پادشاهی تک یاخته.برخی از میکروارگانیسمهایی که قبلاً قارچها یا تک یاختهها در نظر گرفته میشدند به پادشاهی جدیدی منتقل شدهاند کرومیستا(صفحات کرومی). قارچ ها میکروارگانیسم های یوکاریوتی غیر فتوسنتزی (بدون کلروفیل) چند سلولی یا تک سلولی با دیواره سلولی ضخیم هستند. آنها دارای یک هسته با پوشش هسته ای، سیتوپلاسم با اندامک ها، یک غشای سیتوپلاسمی و یک دیواره سلولی سفت و سخت چند لایه متشکل از چندین نوع پلی ساکارید (مانان، گلوکان، سلولز، کیتین) و همچنین پروتئین، لیپیدها و غیره هستند. برخی از قارچ ها تشکیل می شوند. یک کپسول غشای سیتوپلاسمی حاوی گلیکوپروتئینها، فسفولیپیدها و ارگوسترولها (بر خلاف کلسترول، استرول اصلی بافتهای پستانداران) است. بیشتر قارچ ها هوازی اجباری یا اختیاری هستند.
قارچ ها در طبیعت به خصوص در خاک گسترده هستند. برخی از قارچ ها به تولید نان، پنیر، محصولات اسید لاکتیک و الکل کمک می کنند. سایر قارچ ها آنتی بیوتیک های ضد میکروبی (مانند پنی سیلین) و داروهای سرکوب کننده سیستم ایمنی (مانند سیکلوسپورین) تولید می کنند. قارچها توسط ژنتیکشناسان و زیستشناسان مولکولی برای مدلسازی فرآیندهای مختلف استفاده میشوند. قارچ های فیتوپاتوژن آسیب قابل توجهی به کشاورزی وارد می کنند و باعث بیماری های قارچی گیاهان غلات و غلات می شوند. عفونت های ناشی از قارچ ها مایکوز نامیده می شود. قارچ های هیف و مخمری وجود دارد.
قارچ های هیف (کپک) یا هیفومیست ها از نخ های نازکی به ضخامت 2 تا 50 میکرون تشکیل شده اند که هیف نامیده می شوند و به صورت میسلیوم یا میسلیوم (قالب) بافته می شوند. بدن قارچ تالوس نامیده می شود. هیفومیست های دماسیوم (رنگدانه دار - قهوه ای یا سیاه) و هیالین (غیر رنگدانه) وجود دارد. هیف هایی که در بستر ماده غذایی رشد می کنند وظیفه تغذیه قارچ را بر عهده دارند و هیف های رویشی نامیده می شوند. هیف، را-
که بالای سطح زیرلایه قرار می گیرند، هیف های هوایی یا تولید مثلی (مسئول تولید مثل) نامیده می شوند. کلنی ها به دلیل میسلیوم هوایی ظاهری کرکی دارند.
قارچهای پایینتر و بالاتر وجود دارند: هیفهای قارچهای بالاتر با پارتیشنها یا سپتومهایی با سوراخ از هم جدا میشوند. هیف های قارچ های پایینی دارای پارتیشن نیستند و سلول های چند هسته ای هستند که کوئنوسیتیک (از یونانی) نامیده می شوند. کوئنوس- مجرد، مشترک).
قارچ های مخمر (مخمر) عمدتاً توسط سلول های بیضی شکل فردی با قطر 3-15 میکرون نشان داده می شوند و کلنی های آنها بر خلاف قارچ های هیف دارای ظاهر فشرده هستند. با توجه به نوع تولید مثل جنسی، آنها در بین قارچ های بالاتر - آسکومیست ها و بازیدیومیست ها توزیع می شوند. هنگام تولید مثل غیرجنسی، مخمر جوانه می زند یا تقسیم می شود. آنها می توانند شبه هیف ها و میسلیوم های کاذب (سئودومیسلیوم) را به شکل زنجیره ای از سلول های دراز - "سوسیس" تشکیل دهند. قارچ هایی که شبیه مخمر هستند، اما روش تولید مثل جنسی ندارند، مخمر مانند نامیده می شوند. آنها فقط به صورت غیرجنسی تولید مثل می کنند - با جوانه زدن یا شکافت. مفاهیم "قارچ های مخمر مانند" اغلب با مفهوم "مخمر" شناسایی می شوند.
بسیاری از قارچ ها دارای دوشکلی هستند - توانایی رشد هیف (میسلیوم) یا مخمر مانند، بسته به شرایط کشت. در یک ارگانیسم آلوده، آنها به شکل سلول های مخمر مانند (فاز مخمر) رشد می کنند و در محیط های غذایی هیف و میسلیوم تشکیل می دهند. دیمورفیسم با عامل دما همراه است: در دمای اتاق میسلیوم تشکیل می شود و در دمای 37 درجه سانتیگراد (در دمای بدن انسان) سلول های مخمر مانند تشکیل می شود.
قارچ ها به صورت جنسی یا غیرجنسی تولید مثل می کنند. تولیدمثل جنسی قارچ ها با تشکیل گامت ها، هاگ های جنسی و سایر اشکال جنسی اتفاق می افتد. به اشکال جنسی تئومورف می گویند.
تولید مثل غیرجنسی قارچ ها با تشکیل اشکال مربوطه به نام آنامورف اتفاق می افتد. چنین تولید مثلی با جوانه زدن، تکه تکه شدن هیف ها و هاگ های غیرجنسی اتفاق می افتد. اسپورهای درون زا (اسپورانژیوسپورها) در داخل یک ساختار گرد - اسپورانژیوم - بالغ می شوند. هاگ های برون زا (کنیدی) در نوک هیف های میوه دهی تشکیل می شوند که اصطلاحاً کونیدیوفورها نامیده می شوند.
کنیدی های مختلفی وجود دارد. آرتروکونیدیا (آرتروسپورها) یا تالوکونیدیا با جدا شدن یکنواخت و تکه تکه شدن هیفها و بلاستوکونیدیاها در نتیجه جوانه زدن به وجود می آیند. کنیدی های تک سلولی کوچک را میکروکنیدی و کنیدی های چند سلولی بزرگ را ماکروکنیدی می نامند. اشکال غیرجنسی قارچ ها نیز شامل کلامیدوکونیدیا یا کلامیدوسپورها (سلول های استراحت بزرگ با دیواره ضخیم یا مجموعه ای از سلول های کوچک) می شوند.
قارچ های کامل و ناقص وجود دارد. قارچ های کامل روش تولید مثل جنسی دارند. اینها شامل زیگومیست ها هستند (زیگومیکوتا)،آسکومیست ها (آسکومیکوتا)و بازیدیومیست ها (Basidiomycota).قارچ های ناقص فقط تولید مثل غیرجنسی دارند. اینها شامل نوع/گروه مرسوم رسمی قارچ ها - دوترومیست ها است (دیترومایکوتا).
زیگومیست ها متعلق به قارچ های پایینی (میسلیوم غیر سپتات) هستند. آنها شامل نمایندگان جنس هستند Mucor، Rhizopus، Rhizomucor، Absidia، Basidiobolus، Conidiobolus.در خاک و هوا پخش می شود. آنها می توانند باعث زیگومایکوزیس (موکورومیکوز) ریه ها، مغز و سایر اندام های انسان شوند.
در طی تولید مثل غیرجنسی زیگومیست ها، یک اسپورانژیوم روی هیفای باردهی (اسپورانژیوفورها) تشکیل می شود - یک ضخیم شدن کروی با پوسته ای حاوی اسپورانژیوسپورهای متعدد (شکل 2.6، 2.7). تولید مثل جنسی در زیگومیست ها با کمک زیگوسپورها انجام می شود.
آسکومیست ها (قارچ های کیسه دار) دارای میسلیوم سپتات هستند (به جز مخمرهای تک سلولی). آنها نام خود را از اندام اصلی میوه - بورسا یا آسکوس، حاوی 4 یا 8 هاگ جنسی هاپلوئید (آسکوسپور) گرفته اند.
آسکومیست ها شامل نمایندگان منفرد (تلئومورف) جنس هستند آسپرژیلوسو پنی سیلیوماکثر جنس های قارچ آسپرژیلوس، پنی سیلیومآنامورفیک هستند، یعنی آنها فقط بدون درمان تولید مثل می کنند
 برنج. 2.6.قارچ از جنس موکور(نقاشی توسط A.S. Bykov)
برنج. 2.6.قارچ از جنس موکور(نقاشی توسط A.S. Bykov)
 برنج. 2.7.قارچ از جنس ریزوپوس.رشد اسپورانژیوم، اسپورانژیوسپورها و ریزوئیدها
برنج. 2.7.قارچ از جنس ریزوپوس.رشد اسپورانژیوم، اسپورانژیوسپورها و ریزوئیدها
به طور مستقیم با کمک اسپورهای غیرجنسی - کنیدیوم (شکل 2.8، 2.9) و باید بر اساس این ویژگی به عنوان قارچ های ناقص طبقه بندی شوند. در قارچ های جنس آسپرژیلوسدر انتهای هیف های میوه دهی، کنیدیوفورها، ضخیم شدن ها وجود دارد - استریگماتا، فیالیدها، که روی آنها زنجیره های کنیدی ها تشکیل می شود ("قالب آب").
در قارچ های جنس پنی سیلیوم(منگوله) هیفا میوه ای شبیه یک قلم مو است ، زیرا از آن ضخیم می شود (روی کنیدیوفور) که به ساختارهای کوچکتر منشعب می شود - استریگماتا ، فیالیدها ، که روی آنها زنجیره های کنیدی وجود دارد. برخی از گونه های آسپرژیلوس می توانند باعث آسپرژیلوزیس و آفلاتوککسیکوز شوند و پنی سیلیوم می تواند باعث ایجاد پنی سیلیوز شود.
نمایندگان آسکومیست ها تئومورف های این جنس هستند تریکوفیتون، میکروسپوروم، هیستوپلاسما، بلاستومایسس،و همچنین لرزش
 برنج. 2.8.قارچ از جنس پنی سیلیومزنجیره های کندی از فیالیدها گسترش می یابد
برنج. 2.8.قارچ از جنس پنی سیلیومزنجیره های کندی از فیالیدها گسترش می یابد
 برنج. 2.9.قارچ از جنس آسپرژیلوس فومیگاتوس.زنجیره های کندی از فیالیدها گسترش می یابد
برنج. 2.9.قارچ از جنس آسپرژیلوس فومیگاتوس.زنجیره های کندی از فیالیدها گسترش می یابد
بازیدیومیست ها شامل قارچ های کلاهکی هستند. آنها میسلیوم سپتات دارند و با جدا شدن از بازیدیوم - سلول انتهایی میسلیوم، همولوگ با آسک، هاگ های جنسی - بازیدیوسپورها را تشکیل می دهند. بازیدیومیست ها شامل برخی از مخمرها مانند تله مورف ها هستند Cryptococcus neoformans.
دوترومیست ها قارچ های ناقصی هستند (قارچ ناقص،قارچ های آنمورفیک، قارچ های مخروطی). این یک تاکسون مشروط و رسمی از قارچ ها است که قارچ هایی را که تولید مثل جنسی ندارند متحد می کند. اخیراً به جای اصطلاح "دوترومیست ها" اصطلاح "قارچ های میتوسپور" پیشنهاد شده است - قارچ هایی که توسط هاگ های غیرجنسی تولید مثل می کنند. توسط میتوز هنگامی که واقعیت تولید مثل جنسی قارچ های ناقص مشخص شد، آنها به یکی از انواع شناخته شده منتقل می شوند - آسکومیکوتایا Basidiomycotaانتساب نام به یک فرم دورنما. دوترومیست ها دارای میسلیوم سپتات هستند و تنها از طریق تشکیل غیرجنسی کنیدی ها تولید مثل می کنند. دوترومیست ها شامل مخمرهای ناقص (قارچ های مخمر مانند) هستند، به عنوان مثال، برخی از قارچ های جنس کاندیدا،پوست، غشاهای مخاطی و اندام های داخلی را تحت تاثیر قرار می دهد (کاندیدیاز). شکل آنها بیضی شکل به قطر 2-5 میکرون است، با جوانه زدن تقسیم می شوند، به شکل زنجیره ای از سلول های دراز، هیف های کاذب (سئودومیسلیوم) را تشکیل می دهند و گاهی اوقات هیف ایجاد می کنند. برای کاندیدا آلبیکنستشکیل کلامیدوسپورها مشخص است (شکل 2.10). دوترومیست ها همچنین شامل قارچ های دیگری هستند که روش تولید مثل جنسی ندارند و متعلق به جنس هستند. Epidermophyton، Coccidioides، Paracoccidioides، Sporothrix، Aspergillus، Phialophora، Fonsecaea، Exophiala، Cladophialophora، Bipolaris، Exerohilum، Wangiella، Alrernariaو غیره.
 برنج. 2.10.قارچ از جنس کاندیدا آلبیکنس(نقاشی توسط A.S. Bykov)
برنج. 2.10.قارچ از جنس کاندیدا آلبیکنس(نقاشی توسط A.S. Bykov)
2.4. ساختار و طبقه بندی تک یاخته ها
ساده ترین ها متعلق به دامنه هستند یوکاریا،پادشاهی حیوانات (حیوانات)،زیر پادشاهی تک یاخته.اخیراً پیشنهاد شده است که تک یاخته ها را به رتبه پادشاهی اختصاص دهند تک یاخته.
سلول تک یاخته ای توسط یک غشاء (پلیکل) احاطه شده است - آنالوگ غشای سیتوپلاسمی سلول های حیوانی. دارای یک هسته با پوشش هسته ای و هسته، سیتوپلاسم حاوی شبکه آندوپلاسمی، میتوکندری، لیزوزوم و ریبوزوم است. اندازه تک یاخته ها بین 2 تا 100 میکرون است. هنگام رنگ آمیزی طبق رومانوفسکی-گیمسا، هسته تک یاخته قرمز و سیتوپلاسم آبی است. تک یاخته ها با کمک تاژک، مژک یا شبه پودی حرکت می کنند، برخی از آنها واکوئل های گوارشی و انقباضی (دفعی) دارند. آنها می توانند در نتیجه فاگوسیتوز یا تشکیل ساختارهای ویژه تغذیه شوند. بر اساس نوع تغذیه به هتروتروف و اتوتروف تقسیم می شوند. بسیاری از تک یاخته ها (آمیب دیسانتر، ژیاردیا، تریکوموناس، لیشمانیا، بالانتیدیا) می توانند در محیط های غذایی حاوی پروتئین های بومی و اسیدهای آمینه رشد کنند. از کشت های سلولی، جنین مرغ و حیوانات آزمایشگاهی نیز برای پرورش آنها استفاده می شود.
تک یاخته ها به صورت غیرجنسی تولید مثل می کنند - با تقسیم دو یا چندگانه (اسکیزوگونی) و برخی نیز به صورت جنسی (اسپوروژنی). برخی از تک یاخته ها به صورت خارج سلولی تولید مثل می کنند (ژیاردیا)، در حالی که برخی دیگر به صورت داخل سلولی تولید مثل می کنند (پلاسمودیوم، توکسوپلاسما، لیشمانیا). چرخه زندگی تک یاخته ها با مراحلی مشخص می شود - تشکیل مرحله تروفوزوئیت و مرحله کیست. کیست ها مراحل خفته، مقاوم در برابر تغییرات دما و رطوبت هستند. کیست ها به اسید مقاوم هستند سارکوسیستیس، کریپتوسپوریدیومو ایزوسپورا.
پیش از این، تک یاخته هایی که باعث بیماری در انسان می شوند با 4 نوع 1 نشان داده می شدند. Sarcomastigophora، Apicomplexa، Ciliophora، Microspora).این گونه ها اخیراً به تعداد بیشتری طبقه بندی شده اند و پادشاهی های جدیدی ظهور کرده اند - تک یاختهو کرومیستا(جدول 2.2). به یک پادشاهی جدید کرومیستا(chromovics) شامل برخی تک یاخته ها و قارچ ها (بلاستوسیست ها، اوومیست ها و Rhinosporidium seeberi).پادشاهی تک یاختهشامل آمیب ها، تاژکداران، اسپروزوئرها و مژک داران است. آنها به انواع مختلفی تقسیم می شوند که از جمله آنها می توان به آمیب، تاژک دار، اسپروزوئر و مژک دار اشاره کرد.
جدول 2.2.نمایندگان پادشاهی ها تک یاختهو کرومیستا،از اهمیت پزشکی برخوردار است
 1 نوع سارکوماستیگوفورامتشکل از انواع فرعی سارکودیناو ماستیگوفورا.زیرنوع سارکودینا(sarcodaceae) شامل آمیب دیسانتریک و زیرگروه بود ماستیگوفورا(تاژک ها) - تریپانوزوم ها، لیشمانیا، لامبلیا و تریکوموناس. تایپ کنید آپی کمپلکسشامل کلاس اسپروزوآ(اسپور) که شامل پلاسمودیای مالاریا، توکسوپلاسما، کریپتوسپوریدیوم و غیره بود. کیلیوفوراشامل بالانتیدیا، و نوع میکروسپورا- میکروسپوریدیا
1 نوع سارکوماستیگوفورامتشکل از انواع فرعی سارکودیناو ماستیگوفورا.زیرنوع سارکودینا(sarcodaceae) شامل آمیب دیسانتریک و زیرگروه بود ماستیگوفورا(تاژک ها) - تریپانوزوم ها، لیشمانیا، لامبلیا و تریکوموناس. تایپ کنید آپی کمپلکسشامل کلاس اسپروزوآ(اسپور) که شامل پلاسمودیای مالاریا، توکسوپلاسما، کریپتوسپوریدیوم و غیره بود. کیلیوفوراشامل بالانتیدیا، و نوع میکروسپورا- میکروسپوریدیا
انتهای جدول 2.2
 آمیب ها شامل عامل ایجاد کننده آمیبیاز انسانی - اسهال خونی آمیبی است (Entamoeba histolytica)آمیب های آزاد و غیر بیماری زا (آمیب روده و غیره). آمیب ها باینری به صورت غیرجنسی تولید مثل می کنند. چرخه زندگی آنها شامل یک مرحله تروفوزوئیت (یک سلول در حال رشد، متحرک، ضعیف پایدار) و یک مرحله کیست است. تروفوزوئیت ها با کمک شبه پاها حرکت می کنند که مواد مغذی را جذب کرده و در سیتوپلاسم فرو می برند. از جانب
آمیب ها شامل عامل ایجاد کننده آمیبیاز انسانی - اسهال خونی آمیبی است (Entamoeba histolytica)آمیب های آزاد و غیر بیماری زا (آمیب روده و غیره). آمیب ها باینری به صورت غیرجنسی تولید مثل می کنند. چرخه زندگی آنها شامل یک مرحله تروفوزوئیت (یک سلول در حال رشد، متحرک، ضعیف پایدار) و یک مرحله کیست است. تروفوزوئیت ها با کمک شبه پاها حرکت می کنند که مواد مغذی را جذب کرده و در سیتوپلاسم فرو می برند. از جانب
تروفوزوئیت کیستی را تشکیل می دهد که در برابر عوامل خارجی مقاوم است. هنگامی که در روده قرار می گیرد، به یک تروفوزوئیت تبدیل می شود.
تاژک داران با وجود تاژک مشخص می شوند: لیشمانیا دارای یک تاژک، تریکوموناس دارای 4 تاژک آزاد و یک تاژک متصل به یک غشای موجدار کوتاه است. آن ها هستند:
تاژک های خون و بافت ها (لیشمانیا - عوامل ایجاد کننده لیشمانیوز؛ تریپانوزوم ها - عوامل ایجاد کننده بیماری خواب و بیماری شاگاس).
تاژکک های روده (ژیاردیا - عامل ایجاد کننده ژیاردیازیس)؛
تاژک های دستگاه تناسلی ادراری (Trichomonas vaginalis - عامل ایجاد کننده تریکومونیازیس).
مژک دارها با بالانتیدیا نشان داده می شوند که روده بزرگ انسان را تحت تأثیر قرار می دهند (اسهال خونی بالانتیدیازیس). بالانتیدیا دارای مرحله تروفوزوئیت و کیست است. تروفوزوئیت متحرک، دارای مژک های متعدد، نازک تر و کوتاه تر از تاژک است.
2.5. ساختار و طبقه بندی ویروس ها
ویروس ها کوچکترین میکروب های متعلق به پادشاهی هستند ویرا(از لات ویروس- من). ساختار سلولی ندارند و تشکیل می شوند
ساختار ویروسها به دلیل اندازه کوچکشان، با استفاده از میکروسکوپ الکترونی ویریونها و مقاطع بسیار نازک آنها بررسی میشود. اندازه ویروس ها (ویریون ها) مستقیماً با استفاده از میکروسکوپ الکترونی یا به طور غیرمستقیم توسط اولترافیلتراسیون از طریق فیلترهایی با قطر منافذ مشخص یا با اولتراسانتریفیوژ تعیین می شود. اندازه ویروس ها از 15 تا 400 نانومتر است (1 نانومتر برابر با 1/1000 میکرومتر است): ویروس های کوچک که اندازه آنها شبیه به اندازه ریبوزوم است شامل پاروویروس ها و فلج اطفال و بزرگترین آنها ویروس واریولا است (350). نانومتر). ویروس ها از نظر شکل ویریون هایشان متفاوت هستند که به شکل میله ها (ویروس موزاییک تنباکو)، گلوله ها (ویروس هاری)، کره ها (ویروس های فلج اطفال، HIV)، نخ ها (فیلوویروس ها)، اسپرم (بسیاری از باکتریوفاژها) هستند.
ویروس ها با تنوع ساختار و ویژگی های خود تخیل را شگفت زده می کنند. برخلاف ژنوم های سلولی که حاوی DNA دو رشته ای یکنواخت هستند، ژنوم های ویروسی بسیار متنوع هستند. ویروس های DNA و RNA وجود دارند که هاپلوئید هستند، یعنی. یک سری ژن دارند فقط رتروویروس ها دارای ژنوم دیپلوئید هستند. ژنوم ویروس ها شامل 6 تا 200 ژن است و با انواع مختلفی از اسیدهای نوکلئیک نشان داده می شود: دو رشته ای، تک رشته ای، خطی، دایره ای، تکه تکه شده.
در بین ویروسهای RNA تک رشتهای، بین RNA ژنومی به علاوه رشته و RNA منهای رشته (قطب RNA) تمایز قائل میشود. رشته مثبت (رشته مثبت) RNA این ویروس ها، علاوه بر عملکرد ژنومی (ارثی)، عملکرد RNA اطلاعاتی یا پیام رسان (mRNA یا mRNA) را نیز انجام می دهد. این ماتریکسی برای سنتز پروتئین روی ریبوزوم های سلول آلوده است. RNA بعلاوه رشته عفونی است: هنگامی که به سلول های حساس وارد می شود، می تواند یک فرآیند عفونی ایجاد کند.
cess رشته منهای (رشته منفی) ویروس های RNA فقط یک عملکرد ارثی را انجام می دهد. برای سنتز پروتئین، یک رشته مکمل روی رشته منهای RNA سنتز می شود. برخی از ویروس ها دارای ژنوم RNA دوقطبی هستند (محیطاز یونانی آمبی- در هر دو طرف، مكمل مضاعف)، یعنی. شامل بخش های RNA مثبت و منفی است.
ویروس های ساده (به عنوان مثال، ویروس هپاتیت A) و ویروس های پیچیده (به عنوان مثال، ویروس های آنفولانزا، تبخال، ویروس کرونا) وجود دارد.
ویروسهای ساده یا بدون پوشش فقط اسید نوکلئیک دارند که با ساختار پروتئینی به نام کپسید (از لاتین) مرتبط است. capsa- مورد). پروتئین های مرتبط با اسید نوکلئیک به عنوان نوکلئوپروتئین شناخته می شوند و ارتباط پروتئین های کپسید ویروسی با اسید نوکلئیک ویروسی را نوکلئوکپسید می نامند. برخی از ویروسهای ساده میتوانند کریستال ایجاد کنند (به عنوان مثال ویروس تب برفکی).
کپسید شامل زیر واحدهای مورفولوژیکی تکرار شونده - کپسومرها، متشکل از چندین پلی پپتید است. اسید نوکلئیک ویریون به کپسید متصل می شود و یک نوکلئوکپسید تشکیل می دهد. کپسید از اسید نوکلئیک در برابر تخریب محافظت می کند. در ویروس های ساده، کپسید در اتصال (جذب) به سلول میزبان نقش دارد. ویروس های ساده در نتیجه تخریب سلول (لیز) از سلول خارج می شوند.
ویروسهای پیچیده یا پوششدار (شکل 2.11)، علاوه بر کپسید، دارای یک پوشش لیپوپروتئینی مضاعف غشایی (مترادف: سوپر کپسید یا پپلو) هستند که از طریق جوانه زدن ویریون از طریق غشای سلولی، به عنوان مثال، از طریق غشای سلولی به دست میآیند. غشای پلاسما، غشای هسته ای یا غشای شبکه آندوپلاسمی. پاکت ویروس حاوی گلیکوپروتئین است،
یا خارها، پپلومرها. تخریب پوسته با اتر و سایر حلال ها ویروس های پیچیده را غیرفعال می کند. در زیر پوسته برخی از ویروس ها یک پروتئین ماتریکس (پروتئین M) قرار دارد.
ویریون ها دارای تقارن کپسید مارپیچ، ایکوسادرال (مکعبی) یا پیچیده (نوکلئوکپسید) هستند. نوع مارپیچی تقارن به دلیل ساختار مارپیچی نوکلئوکپسید است (به عنوان مثال، در ویروس های آنفولانزا، ویروس کرونا): کپسومرها به صورت مارپیچی همراه با اسید نوکلئیک قرار گرفته اند. نوع تقارن ایکوسادرال به دلیل تشکیل یک جسم توخالی ایزومتریک از کپسید حاوی نوکلئیک اسید ویروسی است (مثلاً در ویروس هرپس).
کپسید و پوسته (سوپر کپسید) از ویریون ها در برابر تأثیرات محیطی محافظت می کند و برهمکنش انتخابی (جذب) پروتئین های گیرنده آنها را با موارد خاصی تعیین می کند.
 برنج. 2.11.ساختار ویروس های پوشیده شده با کپسید ایکوسادرال (a) و مارپیچ (b)
برنج. 2.11.ساختار ویروس های پوشیده شده با کپسید ایکوسادرال (a) و مارپیچ (b)
سلول ها و همچنین خواص آنتی ژنی و ایمنی زایی ویریون ها.
ساختار داخلی ویروس ها هسته نامیده می شود. در آدنوویروس ها، هسته از پروتئین های هیستون مانند مرتبط با DNA، در ریو ویروس ها - از پروتئین های کپسید داخلی تشکیل شده است.
برنده جایزه نوبل D. Baltimore سیستم طبقه بندی بالتیمور را بر اساس مکانیسم سنتز mRNA پیشنهاد کرد. این طبقه بندی ویروس ها را در 7 گروه قرار می دهد (جدول 2.3). کمیته بین المللی طبقه بندی ویروس ها (ICTV)یک سیستم طبقه بندی جهانی را اتخاذ کرد که از دسته بندی های طبقه بندی مانند خانواده (نام به پایان می رسد viridae)زیرخانواده (نام به پایان می رسد virinae)جنس (نام به پایان می رسد ویروس).گونه ویروس مانند باکتری نام دوجمله ای دریافت نکرد. ویروس ها بر اساس نوع اسید نوکلئیک (DNA یا RNA)، ساختار آن و تعداد رشته ها طبقه بندی می شوند. آنها دارای اسیدهای نوکلئیک دو رشته ای یا تک رشته ای هستند. قطبیت اسید نوکلئیک مثبت (+)، منفی (-) یا قطبیت اسید نوکلئیک مخلوط، دوقطبی (+، -)؛ اسید نوکلئیک خطی یا دایره ای؛ اسید نوکلئیک تکه تکه یا غیر تکه تکه شده اندازه و مورفولوژی ویریون ها، تعداد کپسومرها و نوع تقارن نوکلئوکپسید، وجود پوشش (سوپر کپسید)، حساسیت به اتر و دی اکسی کولات، محل تولید مثل در سلول، خواص آنتی ژنی و غیره نیز گرفته شده است. به حساب آوردن.
جدول 2.3.ویروس های مهم پزشکی
 ادامه جدول. 2.3
ادامه جدول. 2.3
 انتهای جدول 2.3
انتهای جدول 2.3
 ویروس ها حیوانات، باکتری ها، قارچ ها و گیاهان را آلوده می کنند. ویروسها بهعنوان عوامل اصلی بیماریهای عفونی انسان، در فرآیندهای سرطانزایی نیز دخیل هستند و میتوانند به روشهای مختلفی از جمله از طریق جفت (ویروس سرخجه، سیتومگالوویروس و غیره) منتقل شوند و بر جنین انسان تأثیر بگذارند. آنها همچنین می توانند منجر به عوارض پس از عفونی شوند - ایجاد میوکاردیت، پانکراتیت، نقص ایمنی و غیره.
ویروس ها حیوانات، باکتری ها، قارچ ها و گیاهان را آلوده می کنند. ویروسها بهعنوان عوامل اصلی بیماریهای عفونی انسان، در فرآیندهای سرطانزایی نیز دخیل هستند و میتوانند به روشهای مختلفی از جمله از طریق جفت (ویروس سرخجه، سیتومگالوویروس و غیره) منتقل شوند و بر جنین انسان تأثیر بگذارند. آنها همچنین می توانند منجر به عوارض پس از عفونی شوند - ایجاد میوکاردیت، پانکراتیت، نقص ایمنی و غیره.
علاوه بر ویروس ها، اشکال حیات غیر سلولی شامل پریون ها و ویروئیدها نیز می شود. ویروئیدها مولکولهای کوچکی از RNA دایرهای و ابرپیچپیچ هستند که حاوی پروتئین نیستند و باعث بیماریهای گیاهی میشوند. پریون های پاتولوژیک ذرات پروتئین عفونی هستند که در نتیجه تغییر در ساختار پروتئین پریون سلولی طبیعی باعث بیماری های ساختاری خاص می شوند. PrP c) که در بدن حیوانات و انسان وجود دارد. PrP باعملکردهای نظارتی را انجام می دهد. این ژن توسط ژن پریون طبیعی (ژن PrP) که روی بازوی کوتاه کروموزوم 20 انسان قرار دارد کدگذاری می شود. بیماری های پریون به صورت آنسفالوپاتی های اسفنجی شکل قابل انتقال (بیماری کروتزفلد-جاکوب، کورو و غیره) بروز می کنند. در این مورد، پروتئین پریون شکل متفاوت و عفونی به دست می آورد که به عنوان نامگذاری شده است PrP sc(sc از اسکرپی- اسکرپی عفونت پریونی گوسفند و بز است). این پروتئین پریون عفونی ظاهری فیبریل دارد و از نظر ساختار سوم یا چهارتایی با پروتئین پریون معمولی متفاوت است.
وظایف خودآمادگی (خودکنترلی)
آ.میکروب های پروکاریوتی را علامت بزنید:
2. ویروس ها.
3. باکتری ها.
4. پریون ها.
ب.به ویژگی های متمایز یک سلول پروکاریوتی توجه کنید:
1. ریبوزوم های 70S.
2. وجود پپتیدوگلیکان در دیواره سلولی.
3. وجود میتوکندری.
4. مجموعه دیپلوئیدی از ژن ها.
که در.اجزای پپتیدوگلیکان را علامت بزنید:
1. اسیدهای تیکوئیک.
2. N-acetylglucosamine.
3. لیپوپلی ساراید.
4. تتراپپتید.
جی.به ویژگی های ساختاری دیواره سلولی باکتری های گرم منفی توجه کنید:
1. مزودیامینوپیملیک اسید.
2. اسیدهای تیکوئیک.
4. پروتئین های پورین.
D.عملکرد هاگ ها در باکتری ها را نام ببرید:
1. حفظ گونه.
2. مقاومت در برابر حرارت.
3. پراکندگی بستر.
4. تولید مثل.
1. ریکتزیا.
2. اکتینومیست ها.
3. اسپیروکت ها.
4. کلامیدیا.
و.ویژگی های اکتینومیست ها را نام ببرید:
1. هاگ های حرارت پذیر دارند.
2. باکتری های گرم مثبت.
3. دیواره سلولی وجود ندارد.
4. شکل پیچیده ای دارند.
ز.ویژگی های اسپیروکت ها را نام ببرید:
1. باکتری های گرم منفی.
2. دارای دستگاه فیبریلاری موتوری هستند.
3. شکل پیچیده ای دارند.
و.تک یاخته هایی را نام ببرید که دارای کمپلکس آپیکال هستند که به آنها اجازه نفوذ به داخل سلول را می دهد:
1. پلاسمودیوم مالاریا.
3. توکسوپلاسما.
4. کریپتوسپوریدیوم.
به.ویژگی بارز ویروس های پیچیده را نام ببرید:
1. دو نوع اسید نوکلئیک.
2. وجود غشای لیپیدی.
3. کپسید دوگانه.
4. وجود پروتئین های غیر ساختاری. L.قارچ های بالاتر را علامت بزنید:
1. موکور.
2. کاندیدا.
3. پنی سیلیوم
4. آسپرژیلوس.
میکروارگانیسم ها (میکروب ها) موجودات تک سلولی کوچکتر از 0.1 میلی متر هستند که با چشم غیر مسلح دیده نمی شوند. اینها شامل باکتری ها، ریزجلبک ها، برخی از قارچ های رشته ای تحتانی، مخمرها و تک یاخته ها هستند (شکل 1). میکروبیولوژی آنها را مطالعه می کند.
برنج. 1. اشیاء میکروبیولوژی.
در شکل 2. می توانید برخی از نمایندگان تک یاخته های تک سلولی را مشاهده کنید. گاهی اوقات اشیاء این علم شامل ابتدایی ترین موجودات روی زمین است - ویروس هایی که ساختار سلولی ندارند و مجتمع هایی از اسیدهای نوکلئیک (مواد ژنتیکی) و پروتئین هستند. اغلب آنها در یک زمینه مطالعه کاملاً جداگانه (ویروس شناسی) جدا می شوند، زیرا میکروبیولوژی بیشتر با هدف مطالعه موجودات تک سلولی میکروسکوپی است.

برنج. 2. نمایندگان منفرد یوکاریوت های تک سلولی (پرتوزوآ).
علوم algology و mycology که به ترتیب جلبک ها و قارچ ها را مطالعه می کنند، رشته های جداگانه ای هستند که در مطالعه اشیاء زنده میکروسکوپی با میکروبیولوژی همپوشانی دارند. باکتری شناسی شاخه واقعی میکروبیولوژی است. این علم منحصراً به مطالعه میکروارگانیسم های پروکاریوتی می پردازد (شکل 3).

برنج. 3. طرح یک سلول پروکاریوتی.
برخلاف یوکاریوت ها که شامل همه موجودات چند سلولی و همچنین تک یاخته ها، جلبک های میکروسکوپی و قارچ ها می شود، پروکاریوت ها هسته تشکیل شده حاوی مواد ژنتیکی و اندامک های واقعی (ساختارهای تخصصی دائمی سلول) ندارند.
پروکاریوت ها شامل باکتری های واقعی و باستانی هستند که طبق طبقه بندی مدرن به عنوان دامنه ها (ابر پادشاهی ها) Archaea و Eubacteria تعیین می شوند (شکل 4).

برنج. 4. حوزه های طبقه بندی بیولوژیکی مدرن.
ویژگی های ساختاری باکتری ها
باکتری ها حلقه مهمی در چرخه مواد در طبیعت هستند؛ آنها بقایای گیاهی و حیوانی را تجزیه می کنند، آب های آلوده به مواد آلی را تمیز می کنند و ترکیبات معدنی را اصلاح می کنند. بدون آنها زندگی روی زمین نمی توانست وجود داشته باشد. این میکروارگانیسم ها در همه جا پراکنده هستند، در خاک، آب، هوا، موجودات جانوری و گیاهی.
باکتری ها در ویژگی های مورفولوژیکی زیر متفاوت هستند:
- شکل سلولی (گرد، میله ای شکل، رشته ای، پیچ خورده، مارپیچی، و همچنین گزینه های انتقالی مختلف و پیکربندی ستاره ای شکل).
- وجود وسایل حرکتی (بی حرکت، تاژک دار، به دلیل ترشح مخاط).
- ارتباط سلول ها با یکدیگر (منزوی شده، به شکل جفت، گرانول، اشکال انشعاب پیوند خورده است).
در میان ساختارهایی که توسط باکتریهای گرد (کوکسی) ایجاد میشوند، سلولهایی وجود دارند که پس از تقسیم به صورت جفت هستند و سپس به شکلهای منفرد (میکروکوکی) شکسته میشوند یا همیشه در کنار هم میمانند (دیپلوکوک). یک ساختار درجه دوم از چهار سلول توسط تتراکوک ها، یک زنجیره توسط استرپتوکوک، یک گرانول 8-64 واحدی توسط سارسینا و خوشه ها توسط استافیلوکوک ها تشکیل می شود.
باکتری های میله ای شکل به دلیل تنوع زیاد در طول (0.1-15 میکرومتر) و ضخامت (0.1-2 میکرومتر) سلول با اشکال مختلف نشان داده می شوند. شکل دومی همچنین به توانایی باکتری ها برای تشکیل هاگ بستگی دارد - ساختارهایی با پوسته ضخیم که به میکروارگانیسم ها اجازه می دهد در شرایط نامطلوب زنده بمانند. سلول هایی که این توانایی را دارند باسیل نامیده می شوند و آنهایی که چنین خاصیتی ندارند به سادگی باکتری میله ای شکل نامیده می شوند.
تغییرات ویژه باکتری های میله ای شکل، فرم های رشته ای (دراز)، زنجیره ها و ساختارهای شاخه ای هستند. دومی توسط اکتینومیست ها در مرحله خاصی از رشد تشکیل می شود. میله های "منحنی" را باکتری های چین دار می نامند که در میان آنها ویبریوها جدا می شوند. اسپیرلا دارای دو خم (15-20 میکرومتر)؛ اسپیروکت هایی که شبیه خطوط مواج هستند. طول سلول آنها به ترتیب 1-3، 15-20 و 20-30 میکرومتر است. در شکل شکل های 5 و 6 اشکال مورفولوژیکی اصلی باکتری ها و همچنین انواع آرایش هاگ را در سلول نشان می دهد.

برنج. 5. اشکال اساسی باکتری ها.

برنج. 6. باکتری با توجه به نوع محل اسپور در سلول. 1، 4 - در مرکز؛ 2، 3، 5 - مکان پایانی؛ 6- از پهلو
ساختارهای سلولی اصلی باکتری ها: نوکلوئید (مواد ژنتیکی)، ریبوزوم های در نظر گرفته شده برای سنتز پروتئین، غشای سیتوپلاسمی (بخشی از غشای سلولی)، که در بسیاری از نمایندگان علاوه بر این از بالا، کپسول و غلاف مخاطی محافظت می شود (شکل 7).

برنج. 7. طرح یک سلول باکتریایی.
بر اساس طبقه بندی باکتری ها، بیش از 20 نوع وجود دارد. به عنوان مثال، بسیار گرما دوست (عاشقان دمای بالا) Aquificae، باکتری های میله ای شکل بی هوازی Bacteroidetes. با این حال، غالب ترین شاخه، که شامل نمایندگان متنوعی است، اکتینوباکتریا است. این شامل بیفیدوباکتری ها، لاکتوباسیل ها و اکتینومیست ها است. منحصر به فرد دومی در توانایی تشکیل میسلیوم در مرحله خاصی از رشد نهفته است.
در اصطلاح رایج به آن میسلیوم می گویند. در واقع، سلول های انشعاب اکتینومیست ها شبیه هیف های قارچی هستند. با وجود این ویژگی، اکتینومیست ها به عنوان باکتری طبقه بندی می شوند، زیرا پروکاریوت هستند. به طور طبیعی، سلول های آنها از نظر ساختار کمتر شبیه قارچ ها هستند.
اکتینومیست ها (شکل 8) باکتری هایی با رشد آهسته هستند و بنابراین توانایی رقابت برای سوبستراهای در دسترس را ندارند. آنها قادر به تجزیه موادی هستند که سایر میکروارگانیسم ها نمی توانند از آنها به عنوان منبع کربن استفاده کنند، به ویژه هیدروکربن های نفتی. بنابراین، اکتینومیست ها به شدت در زمینه بیوتکنولوژی مورد مطالعه قرار می گیرند.
برخی از نمایندگان در مناطقی از میادین نفتی متمرکز می شوند و فیلتر باکتریایی خاصی ایجاد می کنند که از نفوذ هیدروکربن ها به جو جلوگیری می کند. اکتینومیست ها تولیدکنندگان فعال ترکیبات با ارزش عملی هستند: ویتامین ها، اسیدهای چرب، آنتی بیوتیک ها.

برنج. 8. نماینده اکتینومیست نوکاردیا.
قارچ در میکروبیولوژی
هدف میکروبیولوژی فقط قارچ های قالب پایین (به ویژه ریزوپوس، موکور) است. مانند همه قارچ ها، آنها قادر به سنتز مواد نیستند و به یک محیط غذایی نیاز دارند. میسلیوم نمایندگان پایین این پادشاهی بدوی است و با پارتیشن تقسیم نمی شود. جایگاه ویژه ای در تحقیقات میکروبیولوژیکی توسط مخمر اشغال شده است (شکل 9) که با عدم وجود میسلیوم مشخص می شود.

برنج. 9. اشکال کلنی های کشت مخمر روی یک محیط غذایی.
در حال حاضر، دانش زیادی در مورد خواص مفید آنها جمع آوری شده است. با این حال، مخمر به دلیل توانایی آن در سنتز ترکیبات آلی با ارزش عملا مورد مطالعه قرار می گیرد و به طور فعال به عنوان موجودات مدل در آزمایشات ژنتیکی استفاده می شود. از زمان های قدیم، مخمر در فرآیندهای تخمیر استفاده می شده است. متابولیسم در بین نمایندگان مختلف متفاوت است. بنابراین، برخی از مخمرها برای یک فرآیند خاص مناسب تر از بقیه هستند.
به عنوان مثال، Saccharomyces beticus، که در برابر غلظت های بالای الکل مقاوم تر است، برای ایجاد شراب های قوی (تا 24٪) استفاده می شود. در حالی که مخمر S. cerevisiae قادر به تولید غلظت کمتر اتانول است. مخمرها با توجه به حوزه کاربردشان به خوراکی، نانوایی، آبجو، الکل و شراب تقسیم می شوند.
میکروارگانیسم های بیماری زا
میکروارگانیسم های بیماری زا یا بیماری زا در همه جا یافت می شوند. همراه با ویروس های شناخته شده: آنفولانزا، هپاتیت، سرخک، HIV و غیره، میکروارگانیسم های خطرناک ریکتزیا و همچنین استرپتوکوک ها و استافیلوکوک ها هستند که باعث مسمومیت خون می شوند. در میان باکتری های میله ای شکل، پاتوژن های زیادی وجود دارد. به عنوان مثال، دیفتری، سل، تب حصبه (شکل 10). بسیاری از نمایندگان میکروارگانیسم های خطرناک برای انسان در میان تک یاخته ها، به ویژه پلاسمودیوم مالاریا، توکسوپلاسما، لیشمانیا، لامبلیا، تریکوموناس و آمیب های بیماری زا یافت می شوند.

برنج. 10. عکس باکتری باسیلوس آنتراسیس عامل سیاه زخم.
بسیاری از اکتینومیست ها برای انسان و حیوانات خطرناک نیستند. با این حال، بسیاری از نمایندگان بیماری زا در میان مایکوباکتریوم ها یافت می شوند که باعث سل و جذام می شوند. برخی از اکتینومیست ها شروع کننده بیماری مانند اکتینومیکوز هستند که با تشکیل گرانولوم و گاهی اوقات افزایش دمای بدن همراه است. انواع خاصی از قارچ های کپک قادر به تولید مواد سمی برای انسان - مایکوتوکسین ها هستند. به عنوان مثال، برخی از نمایندگان جنس Aspergillus، Fusarium. قارچ های بیماری زا باعث ایجاد گروهی از بیماری ها به نام مایکوز می شوند. بنابراین، کاندیدیازیس یا، به زبان ساده، برفک دهان توسط قارچ های مخمر مانند ایجاد می شود (شکل 11). آنها همیشه در بدن انسان وجود دارند، اما تنها زمانی فعال می شوند که سیستم ایمنی ضعیف شود.

برنج. 11. قارچ کاندیدا عامل ایجاد برفک است.
قارچ ها می توانند انواع ضایعات پوستی، به ویژه انواع گلسنگ ها را ایجاد کنند، به جز هرپس زوستر که توسط ویروس ایجاد می شود. مخمر مالاسزیا، ساکنان دائمی پوست انسان، می تواند باعث کاهش فعالیت سیستم ایمنی بدن شود. بلافاصله برای شستن دست ها عجله نکنید. مخمرها و باکتریهای فرصتطلب در سلامتی خوب عملکرد مهمی را انجام میدهند و از توسعه پاتوژنها جلوگیری میکنند.
ویروس ها به عنوان یک موضوع میکروبیولوژی
ویروس ها ابتدایی ترین موجودات روی زمین هستند. در حالت آزاد، هیچ فرآیند متابولیکی در آنها رخ نمی دهد. ویروس ها تنها زمانی که وارد سلول میزبان می شوند شروع به تکثیر می کنند. در تمام موجودات زنده، ناقل ماده ژنتیکی اسید دئوکسی ریبونوکلئیک (DNA) است. فقط در بین ویروس ها نمایندگانی با یک توالی ژنتیکی مانند اسید ریبونوکلئیک (RNA) وجود دارد.
ویروس ها اغلب به عنوان موجودات زنده واقعی طبقه بندی نمی شوند.
مورفولوژی ویروس ها بسیار متنوع است (شکل 12). به طور معمول، اندازه قطر آنها بین 20-300 نانومتر است.

برنج. 12. تنوع ذرات ویروسی.
طول برخی از نمایندگان به 1-1.5 میکرون می رسد. ساختار ویروس شامل احاطه مواد ژنتیکی با یک چارچوب پروتئینی خاص (کاپسید) است که با اشکال مختلف (مارپیچ، ایکو وجهی، کروی) مشخص می شود. برخی از ویروسها نیز دارای پوششی هستند که از غشای سلول میزبان (سوپر کپسید) تشکیل شده است. به عنوان مثال، (شکل 13) به عنوان عامل بیماری به نام (ایدز) شناخته می شود. حاوی RNA به عنوان ماده ژنتیکی است و بر نوع خاصی از سلول های سیستم ایمنی (لنفوسیت های T کمک کننده) تأثیر می گذارد.

برنج. 13. ساختار ویروس نقص ایمنی انسانی.
باکتری ها قدیمی ترین گروه از موجودات موجود در زمین هستند. اولین باکتری احتمالاً بیش از 3.5 میلیارد سال پیش ظاهر شد و تقریباً برای یک میلیارد سال آنها تنها موجودات زنده روی سیاره ما بودند. از آنجایی که اینها اولین نمایندگان طبیعت زنده بودند، بدن آنها ساختار بدوی داشت.
با گذشت زمان، ساختار آنها پیچیده تر شد، اما تا به امروز باکتری ها به عنوان ابتدایی ترین موجودات تک سلولی در نظر گرفته می شوند. جالب است که برخی از باکتری ها هنوز ویژگی های اولیه اجداد باستانی خود را حفظ کرده اند. این در باکتریهایی که در چشمههای گوگردی گرم و گلهای بدون اکسیژن در کف مخازن زندگی میکنند، مشاهده میشود.
بیشتر باکتری ها بی رنگ هستند. فقط تعداد کمی بنفش یا سبز هستند. اما کلنی های بسیاری از باکتری ها رنگ روشنی دارند که در اثر انتشار یک ماده رنگی در محیط یا رنگدانه شدن سلول ها ایجاد می شود.
کاشف دنیای باکتری ها Antony Leeuwenhoek، طبیعت شناس هلندی قرن هفدهم بود، که برای اولین بار یک میکروسکوپ بزرگنمایی عالی ایجاد کرد که اجسام را 160-270 برابر بزرگنمایی می کرد.
باکتری ها به عنوان پروکاریوت ها طبقه بندی می شوند و به یک پادشاهی جداگانه - باکتری ها طبقه بندی می شوند.
فرم بدن
باکتری ها موجودات متعدد و متنوعی هستند. آنها از نظر شکل متفاوت هستند.
| نام باکتری | شکل باکتری | تصویر باکتری |
| کوکسی | توپی شکل | |
| باسیلوس |  | میله ای شکل |
| ویبریو | کاما شکل | |
| اسپیریلوم |  | مارپیچ |
| استرپتوکوک ها |  | زنجیره ای از کوکسی ها |
| استافیلوکوک |  | خوشه های کوکسی |
| دیپلوکوکوس | دو باکتری گرد محصور در یک کپسول مخاطی |
روش های حمل و نقل
در بین باکتری ها اشکال متحرک و بی حرکت وجود دارد. متحرک ها به دلیل انقباضات موج مانند یا با کمک تاژک ها (نخ های مارپیچ پیچ خورده) حرکت می کنند که از پروتئین خاصی به نام فلاژلین تشکیل شده است. ممکن است یک یا چند تاژک وجود داشته باشد. در برخی از باکتری ها آنها در یک انتهای سلول قرار دارند، در برخی دیگر - در دو یا در کل سطح.

اما حرکت در بسیاری از باکتری های دیگر که فاقد تاژک هستند نیز ذاتی است. بنابراین، باکتری هایی که در خارج با مخاط پوشانده شده اند، می توانند حرکت کنند.
برخی از باکتری های آب و خاک فاقد تاژک دارای واکوئل های گازی در سیتوپلاسم هستند. ممکن است 40-60 واکوئل در یک سلول وجود داشته باشد. هر یک از آنها با گاز (احتمالاً نیتروژن) پر شده است. با تنظیم مقدار گاز در واکوئل ها، باکتری های آبزی می توانند در ستون آب فرو رفته یا به سطح آن بروند و باکتری های خاک می توانند در مویرگ های خاک حرکت کنند.
زیستگاه
به دلیل سازمان دهی ساده و بی تکلفی، باکتری ها در طبیعت گسترده هستند. باکتری ها در همه جا یافت می شوند: حتی در یک قطره از خالص ترین آب چشمه، در دانه های خاک، در هوا، روی سنگ ها، در برف های قطبی، شن های بیابان، در کف اقیانوس ها، در نفت استخراج شده از اعماق زیاد، و حتی در آب چشمه های آب گرم با دمای حدود 80 درجه سانتیگراد. آنها روی گیاهان، میوه ها، حیوانات مختلف و در انسان در روده ها، حفره دهان، اندام ها و روی سطح بدن زندگی می کنند.
باکتری ها کوچک ترین و پرتعدادترین موجودات زنده هستند. به دلیل اندازه کوچک خود به راحتی در هر شکاف، شکاف یا منافذی نفوذ می کنند. بسیار مقاوم و سازگار با شرایط مختلف زندگی. آنها خشک شدن، سرمای شدید و گرم شدن تا دمای 90 درجه سانتیگراد را بدون از دست دادن قابلیت حیات خود تحمل می کنند.
عملاً هیچ مکانی روی زمین وجود ندارد که باکتری در آن یافت نشود، اما در مقادیر متفاوت. شرایط زندگی باکتری ها متفاوت است. برخی از آنها به اکسیژن اتمسفر نیاز دارند، برخی دیگر به آن نیاز ندارند و می توانند در محیطی بدون اکسیژن زندگی کنند.
در هوا: باکتری ها تا 30 کیلومتری جو بالا می روند. و بیشتر.
به خصوص تعداد زیادی از آنها در خاک وجود دارد. 1 گرم خاک می تواند حاوی صدها میلیون باکتری باشد.
در آب: در لایه های سطحی آب در مخازن باز. باکتری های مفید آبزی، بقایای آلی را معدنی می کنند.
در موجودات زنده: باکتری های بیماری زا از محیط خارجی وارد بدن می شوند، اما فقط در شرایط مساعد باعث بیماری می شوند. همزیستی در اندام های گوارشی زندگی می کند و به تجزیه و جذب غذا و سنتز ویتامین ها کمک می کند.
ساختار خارجی
سلول باکتری با یک پوسته متراکم ویژه پوشیده شده است - یک دیواره سلولی که عملکردهای محافظتی و حمایتی را انجام می دهد و همچنین به باکتری شکل دائمی و مشخصه می دهد. دیواره سلولی یک باکتری شبیه دیواره سلول گیاهی است. نفوذپذیر است: از طریق آن مواد مغذی آزادانه وارد سلول می شوند و محصولات متابولیک به محیط خارج می شوند. اغلب، باکتری ها یک لایه محافظ اضافی از مخاط در بالای دیواره سلولی تولید می کنند - یک کپسول. ضخامت کپسول می تواند چندین برابر بیشتر از قطر خود سلول باشد، اما همچنین می تواند بسیار کوچک باشد. کپسول جزء ضروری سلول نیست، بسته به شرایطی که باکتری ها در آن قرار می گیرند تشکیل می شود. از باکتری ها در برابر خشک شدن محافظت می کند.
روی سطح برخی از باکتری ها تاژک های بلند (یک، دو یا چند) یا پرزهای نازک کوتاه وجود دارد. طول تاژک می تواند چندین برابر بزرگتر از اندازه بدن باکتری باشد. باکتری ها با کمک تاژک ها و پرزها حرکت می کنند.

ساختار داخلی
در داخل سلول باکتری سیتوپلاسم متراکم و بی حرکت وجود دارد. دارای ساختار لایه ای است ، واکوئل وجود ندارد ، بنابراین پروتئین های مختلف (آنزیم ها) و مواد مغذی ذخیره در ماده خود سیتوپلاسم قرار دارند. سلول های باکتریایی هسته ندارند. یک ماده حامل اطلاعات ارثی در قسمت مرکزی سلول آنها متمرکز شده است. باکتری، - اسید نوکلئیک - DNA. اما این ماده به صورت هسته تشکیل نمی شود.
سازماندهی داخلی یک سلول باکتری پیچیده است و ویژگی های خاص خود را دارد. سیتوپلاسم توسط غشای سیتوپلاسمی از دیواره سلولی جدا می شود. در سیتوپلاسم یک ماده اصلی یا ماتریس، ریبوزوم ها و تعداد کمی از ساختارهای غشایی وجود دارد که عملکردهای مختلفی را انجام می دهند (آنالوگ های میتوکندری، شبکه آندوپلاسمی، دستگاه گلژی). سیتوپلاسم سلول های باکتریایی اغلب حاوی گرانول هایی با اشکال و اندازه های مختلف است. گرانول ها ممکن است از ترکیباتی تشکیل شده باشند که به عنوان منبع انرژی و کربن عمل می کنند. قطرات چربی نیز در سلول باکتری یافت می شود.
در قسمت مرکزی سلول، ماده هسته ای موضعی است - DNA، که با یک غشاء از سیتوپلاسم جدا نمی شود. این یک آنالوگ هسته است - یک نوکلوئید. نوکلوئید غشاء، هسته یا مجموعه ای از کروموزوم ها ندارد.
روش های غذا خوردن
باکتری ها روش های تغذیه متفاوتی دارند. در میان آنها اتوتروف و هتروتروف وجود دارد. اتوتروف ها موجوداتی هستند که به طور مستقل قادر به تولید مواد آلی برای تغذیه خود هستند.
گیاهان به نیتروژن نیاز دارند، اما نمی توانند نیتروژن را از هوا جذب کنند. برخی از باکتری ها مولکول های نیتروژن موجود در هوا را با مولکول های دیگر ترکیب می کنند و در نتیجه موادی را به وجود می آورند که در دسترس گیاهان هستند.
این باکتریها در سلولهای ریشههای جوان مستقر میشوند که منجر به ایجاد ضخامتهایی روی ریشهها میشود که ندول نامیده میشوند. چنین گره هایی روی ریشه گیاهان خانواده حبوبات و برخی گیاهان دیگر ایجاد می شود.
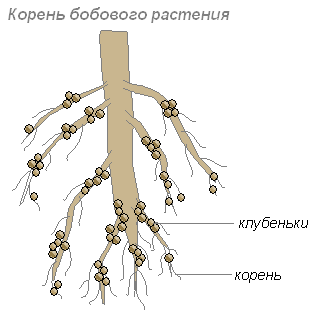
ریشه ها کربوهیدرات ها را برای باکتری ها تامین می کنند و باکتری های ریشه مواد نیتروژن دار را فراهم می کنند که می تواند توسط گیاه جذب شود. زندگی مشترک آنها برای یکدیگر سودمند است.
ریشه گیاهان مواد آلی زیادی (قند، اسیدهای آمینه و غیره) ترشح می کند که باکتری ها از آنها تغذیه می کنند. بنابراین، به خصوص بسیاری از باکتری ها در لایه خاک اطراف ریشه ها مستقر می شوند. این باکتری ها بقایای گیاه مرده را به مواد در دسترس گیاه تبدیل می کنند. این لایه خاک را ریزوسفر می نامند.
چندین فرضیه در مورد نفوذ باکتری های گره به بافت ریشه وجود دارد:
- از طریق آسیب به بافت اپیدرم و قشر مغز؛
- از طریق موهای ریشه؛
- فقط از طریق غشای سلول جوان؛
- به لطف باکتری های همراه که آنزیم های پکتینولیتیک تولید می کنند.
- به دلیل تحریک سنتز B-indoleacetic اسید از تریپتوفان، همیشه در ترشحات ریشه گیاه وجود دارد.
فرآیند ورود باکتری ندول به بافت ریشه شامل دو مرحله است:
- عفونت موهای ریشه؛
- فرآیند تشکیل ندول
در بیشتر موارد، سلول مهاجم به طور فعال تکثیر می شود، به اصطلاح رشته های عفونت را تشکیل می دهد و به شکل چنین رشته هایی به داخل بافت گیاه حرکت می کند. باکتری های ندول که از نخ عفونت بیرون می آیند به تکثیر در بافت میزبان ادامه می دهند.

سلول های گیاهی پر از سلول های تکثیر سریع باکتری های گره شروع به تقسیم سریع می کنند. اتصال یک گره جوان با ریشه یک گیاه حبوبات به لطف بسته های فیبری عروقی انجام می شود. در طول دوره عملکرد، گره ها معمولا متراکم هستند. تا زمانی که فعالیت بهینه رخ می دهد، گره ها رنگ صورتی به خود می گیرند (به لطف رنگدانه لگهموگلوبین). فقط آن دسته از باکتری هایی که حاوی لگهموگلوبین هستند قادر به تثبیت نیتروژن هستند.
باکتری های ندول ده ها و صدها کیلوگرم کود نیتروژن در هر هکتار خاک ایجاد می کنند.
متابولیسم
باکتری ها از نظر متابولیسم با یکدیگر متفاوت هستند. برای برخی با مشارکت اکسیژن اتفاق می افتد، برای برخی دیگر - بدون آن.
بیشتر باکتری ها از مواد آلی آماده تغذیه می کنند. فقط تعداد کمی از آنها (سبز آبی یا سیانوباکتری ها) قادر به ایجاد مواد آلی از مواد معدنی هستند. آنها نقش مهمی در تجمع اکسیژن در جو زمین داشتند.
باکتری ها مواد را از بیرون جذب می کنند، مولکول های خود را تکه تکه می کنند، پوسته خود را از این قسمت ها جمع می کنند و محتویات آنها را دوباره پر می کنند (به این ترتیب رشد می کنند) و مولکول های غیر ضروری را بیرون می اندازند. پوسته و غشای باکتری به آن اجازه می دهد فقط مواد لازم را جذب کند.

اگر پوسته و غشای یک باکتری کاملاً غیر قابل نفوذ بود، هیچ ماده ای وارد سلول نمی شد. اگر آنها به همه مواد نفوذ کنند، محتویات سلول با محیط - محلولی که باکتری در آن زندگی می کند - مخلوط می شود. برای زنده ماندن، باکتری ها به پوسته ای نیاز دارند که اجازه عبور مواد لازم را بدهد، اما نه مواد غیر ضروری.
این باکتری مواد مغذی واقع در نزدیکی خود را جذب می کند. بعد چه اتفاقی می افتد؟ اگر بتواند به طور مستقل حرکت کند (با حرکت دادن تاژک یا عقب راندن مخاط)، آنگاه حرکت می کند تا زمانی که مواد لازم را پیدا کند.
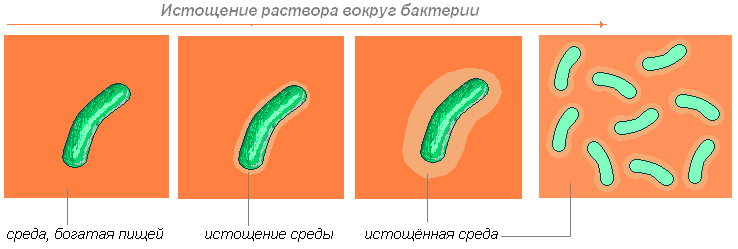
اگر نتواند حرکت کند، صبر می کند تا انتشار (قابلیت نفوذ مولکول های یک ماده به ضخامت مولکول های ماده دیگر) مولکول های لازم را به آن بیاورد.
باکتری ها همراه با سایر گروه های میکروارگانیسم ها، کارهای شیمیایی عظیمی را انجام می دهند. آنها با تبدیل ترکیبات مختلف انرژی و مواد مغذی لازم برای زندگی خود را دریافت می کنند. فرآیندهای متابولیک، روش های به دست آوردن انرژی و نیاز به مواد برای ساختن مواد بدن آنها در باکتری ها متنوع است.
سایر باکتری ها تمام نیازهای خود را به کربن لازم برای سنتز مواد آلی در بدن با هزینه ترکیبات معدنی برآورده می کنند. به آنها اتوتروف می گویند. باکتری های اتوتروف قادر به سنتز مواد آلی از غیر آلی هستند. از جمله آنها عبارتند از:
شیمی سنتز
استفاده از انرژی تابشی مهم ترین، اما نه تنها راه برای ایجاد مواد آلی از دی اکسید کربن و آب است. باکتری هایی شناخته شده اند که از نور خورشید به عنوان منبع انرژی برای چنین سنتزی استفاده نمی کنند، بلکه از انرژی پیوندهای شیمیایی که در سلول های موجودات زنده در طی اکسیداسیون برخی از ترکیبات معدنی - سولفید هیدروژن، گوگرد، آمونیاک، هیدروژن، اسید نیتریک، ترکیبات آهنی رخ می دهد، استفاده می کنند. آهن و منگنز آنها از مواد آلی تشکیل شده با استفاده از این انرژی شیمیایی برای ساخت سلول های بدن خود استفاده می کنند. بنابراین، این فرآیند کموسنتز نامیده می شود.
مهم ترین گروه از میکروارگانیسم های شیمی سنتزی، باکتری های نیتریفیک کننده هستند. این باکتری ها در خاک زندگی می کنند و آمونیاک تشکیل شده در طی تجزیه باقی مانده های آلی به اسید نیتریک را اکسید می کنند. دومی با ترکیبات معدنی خاک واکنش می دهد و به نمک اسید نیتریک تبدیل می شود. این فرآیند در دو مرحله انجام می شود.
باکتری های آهن آهن آهن را به آهن اکسید تبدیل می کنند. هیدروکسید آهن حاصل ته نشین می شود و به اصطلاح سنگ آهن باتلاقی را تشکیل می دهد.
برخی از میکروارگانیسم ها به دلیل اکسیداسیون هیدروژن مولکولی وجود دارند و در نتیجه یک روش تغذیه اتوتروف را ارائه می دهند.
یکی از ویژگیهای باکتریهای هیدروژنی، توانایی تغییر سبک زندگی هتروتروف با ترکیبات آلی و عدم وجود هیدروژن است.
بنابراین، شیمیواتوتروف ها اتوتروف های معمولی هستند، زیرا آنها به طور مستقل ترکیبات آلی لازم را از مواد معدنی سنتز می کنند و آنها را به صورت آماده از موجودات دیگر مانند هتروتروف ها نمی گیرند. باکتری های شیمی اتوتروف با گیاهان فوتوتروف در استقلال کامل آنها از نور به عنوان منبع انرژی تفاوت دارند.
فتوسنتز باکتریایی
برخی از باکتری های گوگرد حاوی رنگدانه (بنفش، سبز)، حاوی رنگدانه های خاص - باکتری کلروفیل ها، قادر به جذب انرژی خورشیدی هستند که با کمک آنها سولفید هیدروژن در بدن آنها شکسته شده و اتم های هیدروژن را برای بازیابی ترکیبات مربوطه آزاد می کند. این فرآیند شباهت زیادی با فتوسنتز دارد و تنها از این جهت متفاوت است که در باکتری های بنفش و سبز، دهنده هیدروژن سولفید هیدروژن (گاهی اوقات اسیدهای کربوکسیلیک) و در گیاهان سبز آب است. در هر دوی آنها جداسازی و انتقال هیدروژن به دلیل انرژی پرتوهای جذب شده خورشید انجام می شود.
این فتوسنتز باکتریایی که بدون آزاد شدن اکسیژن اتفاق می افتد، کاهش نور نامیده می شود. کاهش نور دی اکسید کربن با انتقال هیدروژن نه از آب، بلکه از سولفید هیدروژن همراه است:
6СО 2 +12Н 2 S+hv → С6Н 12 О 6 +12S=6Н 2 О
اهمیت بیولوژیکی شیمی سنتز و فتوسنتز باکتریایی در مقیاس سیاره ای نسبتاً کم است. تنها باکتری های شیمی سنتزی نقش مهمی در روند چرخه گوگرد در طبیعت دارند. گوگرد که توسط گیاهان سبز به شکل نمک های اسید سولفوریک جذب می شود، کاهش یافته و بخشی از مولکول های پروتئین می شود. علاوه بر این، هنگامی که بقایای گیاهی و جانوری مرده توسط باکتری های پوسیده از بین می روند، گوگرد به شکل سولفید هیدروژن آزاد می شود که توسط باکتری های گوگرد به گوگرد آزاد (یا اسید سولفوریک) اکسید می شود و سولفیت هایی را در خاک تشکیل می دهد که در دسترس گیاهان است. باکتری های شیمیایی و فوتو اتوتروفیک در چرخه نیتروژن و گوگرد ضروری هستند.
هاگ زایی
اسپورها در داخل سلول باکتری تشکیل می شوند. در طی فرآیند اسپورزایی، سلول باکتری تحت تعدادی فرآیند بیوشیمیایی قرار می گیرد. مقدار آب آزاد موجود در آن کاهش و فعالیت آنزیمی کاهش می یابد. این امر مقاومت اسپورها را در برابر شرایط نامساعد محیطی (دمای بالا، غلظت نمک زیاد، خشک شدن و غیره) تضمین می کند. اسپورزایی فقط برای گروه کوچکی از باکتری ها مشخص است.

هاگ ها یک مرحله اختیاری در چرخه زندگی باکتری ها هستند. اسپورزایی فقط با کمبود مواد مغذی یا تجمع محصولات متابولیک شروع می شود. باکتری ها به شکل هاگ می توانند برای مدت طولانی غیر فعال بمانند. هاگ های باکتری می توانند در برابر جوشاندن طولانی مدت و انجماد بسیار طولانی مقاومت کنند. هنگامی که شرایط مساعد رخ می دهد، هاگ جوانه می زند و زنده می شود. اسپورهای باکتریایی سازگاری برای زنده ماندن در شرایط نامساعد هستند.
تولید مثل
باکتری ها با تقسیم یک سلول به دو تکثیر می شوند. با رسیدن به اندازه معین، باکتری به دو باکتری یکسان تقسیم می شود. سپس هر یک از آنها شروع به تغذیه می کنند، رشد می کنند، تقسیم می شوند و غیره.

پس از طویل شدن سلول، سپتوم عرضی به تدریج تشکیل می شود و سپس سلول های دختر جدا می شوند. در بسیاری از باکتری ها، تحت شرایط خاصی، پس از تقسیم، سلول ها در گروه های مشخصه به هم متصل می مانند. در این حالت بسته به جهت صفحه تقسیم و تعداد تقسیمات، اشکال مختلفی به وجود می آید. تولید مثل توسط جوانه زدن به عنوان یک استثنا در باکتری ها اتفاق می افتد.
در شرایط مساعد، تقسیم سلولی در بسیاری از باکتری ها هر 20-30 دقیقه اتفاق می افتد. با چنین تولید مثل سریع، فرزندان یک باکتری در 5 روز می توانند توده ای را تشکیل دهند که می تواند تمام دریاها و اقیانوس ها را پر کند. یک محاسبه ساده نشان می دهد که 72 نسل (720,000,000,000,000,000,000 سلول) می تواند در روز تشکیل شود. در صورت تبدیل به وزن - 4720 تن. با این حال، در طبیعت این اتفاق نمی افتد، زیرا اکثر باکتری ها به سرعت تحت تأثیر نور خورشید، خشک شدن، کمبود غذا، گرم شدن تا 65-100 درجه سانتیگراد، در نتیجه مبارزه بین گونه ها و غیره می میرند.
باکتری (1) با جذب غذای کافی، اندازه (2) افزایش می یابد و شروع به آماده شدن برای تولید مثل (تقسیم سلولی) می کند. DNA آن (در یک باکتری، مولکول DNA در یک حلقه بسته است) دو برابر می شود (باکتری یک کپی از این مولکول تولید می کند). هر دو مولکول DNA (3،4) خود را به دیواره باکتری متصل مییابند و با طولانی شدن باکتری، از هم دور میشوند (5،6). ابتدا نوکلئوتید تقسیم می شود، سپس سیتوپلاسم.

پس از واگرایی دو مولکول DNA، انقباض بر روی باکتری ظاهر می شود که به تدریج بدن باکتری را به دو قسمت تقسیم می کند که هر کدام حاوی یک مولکول DNA است (7).
این اتفاق می افتد (در Bacillus subtilis) که دو باکتری به هم می چسبند و پلی بین آنها ایجاد می شود (1،2).

جامپر DNA را از یک باکتری به باکتری دیگر منتقل می کند (3). هنگامی که در یک باکتری، مولکول های DNA در هم تنیده می شوند، در برخی مکان ها به هم می چسبند (4)، و سپس بخش هایی را مبادله می کنند (5).
نقش باکتری ها در طبیعت
مدار
باکتری ها مهمترین حلقه در چرخه کلی مواد در طبیعت هستند. گیاهان مواد آلی پیچیده ای را از دی اکسید کربن، آب و نمک های معدنی در خاک ایجاد می کنند. این مواد با قارچ های مرده، گیاهان و اجساد حیوانات به خاک باز می گردند. باکتری ها مواد پیچیده را به مواد ساده تجزیه می کنند و سپس توسط گیاهان استفاده می شود.
باکتری ها مواد آلی پیچیده گیاهان مرده و اجساد حیوانات، دفع موجودات زنده و مواد زائد مختلف را از بین می برند. با تغذیه از این مواد آلی، باکتری های ساپروفیت پوسیدگی آنها را به هوموس تبدیل می کنند. اینها نوعی نظم دهنده سیاره ما هستند. بنابراین، باکتری ها به طور فعال در چرخه مواد در طبیعت شرکت می کنند.
تشکیل خاک
از آنجایی که باکتری ها تقریباً در همه جا پراکنده شده اند و به تعداد زیادی رخ می دهند، تا حد زیادی فرآیندهای مختلفی را تعیین می کنند که در طبیعت اتفاق می افتد. در پاییز، برگ درختان و درختچه ها می ریزند، شاخه های علف های روی زمین می میرند، شاخه های قدیمی می ریزند و هر از گاهی تنه درختان کهنسال می ریزند. همه اینها به تدریج به هوموس تبدیل می شود. در 1 سانتی متر مکعب. لایه سطحی خاک جنگل حاوی صدها میلیون باکتری ساپروفیت خاک از چندین گونه است. این باکتری ها هوموس را به مواد معدنی مختلفی تبدیل می کنند که می تواند توسط ریشه گیاهان از خاک جذب شود.
برخی از باکتری های خاک قادر به جذب نیتروژن از هوا هستند و از آن در فرآیندهای حیاتی استفاده می کنند. این باکتری های تثبیت کننده نیتروژن به طور مستقل زندگی می کنند یا در ریشه گیاهان حبوبات مستقر می شوند. این باکتری ها با نفوذ به ریشه حبوبات باعث رشد سلول های ریشه و ایجاد گره روی آنها می شوند.
این باکتری ها ترکیبات نیتروژنی تولید می کنند که گیاهان از آنها استفاده می کنند. باکتری ها کربوهیدرات ها و نمک های معدنی را از گیاهان دریافت می کنند. بنابراین، رابطه نزدیکی بین گیاه حبوبات و باکتری ندول وجود دارد که هم برای یکی و هم برای ارگانیسم دیگر مفید است. به این پدیده همزیستی می گویند.
به لطف همزیستی با باکتری های گره، گیاهان حبوبات خاک را با نیتروژن غنی می کنند و به افزایش عملکرد کمک می کنند.
پراکندگی در طبیعت
میکروارگانیسم ها در همه جا وجود دارند. تنها استثناها دهانه های آتشفشان های فعال و مناطق کوچک در کانون بمب های اتمی منفجر شده هستند. نه دمای پایین قطب جنوب، نه جریان های جوشان آبفشان ها، نه محلول های نمک اشباع در استخرهای نمک، نه تابش شدید قله های کوه ها، و نه تابش شدید راکتورهای هسته ای در وجود و توسعه میکرو فلورا دخالت نمی کند. همه موجودات زنده دائماً با میکروارگانیسم ها در تعامل هستند و اغلب نه تنها مخزن آنها، بلکه توزیع کننده آنها نیز هستند. میکروارگانیسم ها بومی سیاره ما هستند و به طور فعال باورنکردنی ترین بسترهای طبیعی را کاوش می کنند.
میکرو فلور خاک
تعداد باکتری ها در خاک بسیار زیاد است - صدها میلیون و میلیاردها نفر در هر گرم. تعداد آنها در خاک بسیار بیشتر از آب و هوا است. تعداد کل باکتری ها در خاک تغییر می کند. تعداد باکتری ها به نوع خاک، وضعیت آنها و عمق لایه ها بستگی دارد.
بر روی سطح ذرات خاک، میکروارگانیسم ها در میکروکلونی های کوچک (هر کدام 20-100 سلول) قرار دارند. آنها اغلب در ضخامت لخته های مواد آلی، روی ریشه های گیاهان زنده و در حال مرگ، در مویرگ های نازک و توده های درونی ایجاد می شوند.
میکرو فلور خاک بسیار متنوع است. در اینجا گروه های فیزیولوژیکی مختلفی از باکتری ها وجود دارد: باکتری های گندیده، باکتری های نیتروژن، باکتری های تثبیت کننده نیتروژن، باکتری های گوگردی و غیره که در میان آنها هوازی و بی هوازی، اشکال هاگ و غیر اسپور وجود دارد. میکرو فلور یکی از عوامل تشکیل خاک است.
منطقه توسعه میکروارگانیسم ها در خاک منطقه مجاور ریشه گیاهان زنده است. به آن ریزوسفر و به مجموع میکروارگانیسم های موجود در آن میکرو فلور ریزوسفر می گویند.
میکرو فلور مخازن
آب یک محیط طبیعی است که در آن میکروارگانیسم ها در تعداد زیادی رشد می کنند. قسمت عمده آنها از خاک وارد آب می شود. عاملی که تعداد باکتری ها در آب و وجود مواد مغذی در آن را تعیین می کند. تمیزترین آب ها از چاه ها و چشمه های آرتزین است. مخازن باز و رودخانه ها از نظر باکتری بسیار غنی هستند. بیشترین تعداد باکتری در لایه های سطحی آب، نزدیک به ساحل یافت می شود. با دور شدن از ساحل و افزایش عمق، تعداد باکتری ها کاهش می یابد.
آب تمیز حاوی 100-200 باکتری در هر میلی لیتر است و آب آلوده حاوی 100-300 هزار یا بیشتر است. باکتری های زیادی در لجن پایین، به خصوص در لایه سطحی، جایی که باکتری ها یک لایه تشکیل می دهند، وجود دارد. این فیلم حاوی مقدار زیادی باکتری های گوگرد و آهن است که سولفید هیدروژن را به اسید سولفوریک اکسید می کند و در نتیجه از مرگ ماهی ها جلوگیری می کند. در گل و لای شکل های هاگ دار بیشتری وجود دارد، در حالی که اشکال غیر هاگ دار در آب غالب است.
از نظر ترکیب گونه، میکرو فلور آب مشابه میکرو فلور خاک است، اما اشکال خاصی نیز وجود دارد. میکروارگانیسم ها با از بین بردن زباله های مختلفی که وارد آب می شوند، به تدریج تصفیه بیولوژیکی آب را انجام می دهند.
میکرو فلور هوا
تعداد میکرو فلور هوا کمتر از میکرو فلور خاک و آب است. باکتری ها همراه با گرد و غبار به هوا می روند، می توانند مدتی در آنجا باقی بمانند و سپس در سطح زمین مستقر شوند و در اثر کمبود تغذیه یا تحت تأثیر اشعه ماوراء بنفش می میرند. تعداد میکروارگانیسم های موجود در هوا به منطقه جغرافیایی، زمین، زمان سال، آلودگی گرد و غبار و غیره بستگی دارد. هر ذره گرد و غبار حامل میکروارگانیسم ها است. بیشتر باکتری ها در هوای بالای شرکت های صنعتی هستند. هوای روستاها تمیزتر است. پاک ترین هوا در جنگل ها، کوه ها و مناطق برفی است. لایه های بالایی هوا حاوی میکروب های کمتری است. میکرو فلور هوا حاوی بسیاری از باکتریهای رنگدانهدار و حاوی اسپور است که نسبت به سایرین در برابر اشعه ماوراء بنفش مقاومتر هستند.
میکرو فلور بدن انسان
بدن انسان، حتی یک بدن کاملا سالم، همیشه حامل میکرو فلور است. هنگامی که بدن انسان با هوا و خاک تماس پیدا می کند، میکروارگانیسم های مختلف از جمله میکروارگانیسم های بیماری زا (باسیل های کزاز، گانگرن گازی و غیره) روی لباس و پوست می نشینند. اغلب قسمت هایی از بدن انسان آلوده است. E. coli و استافیلوکوک در دست یافت می شود. بیش از 100 نوع میکروب در حفره دهان وجود دارد. دهان با دما، رطوبت و بقایای مواد مغذی، محیطی عالی برای رشد میکروارگانیسم ها است.
معده یک واکنش اسیدی دارد، بنابراین اکثر میکروارگانیسم های موجود در آن می میرند. با شروع از روده کوچک، واکنش قلیایی می شود، یعنی. برای میکروب ها مناسب است. میکرو فلور در روده بزرگ بسیار متنوع است. هر فرد بالغ روزانه حدود 18 میلیارد باکتری را از طریق مدفوع دفع می کند. افراد بیشتر از مردم در جهان است.
اندام های داخلی که به محیط خارجی (مغز، قلب، کبد، مثانه و غیره) متصل نیستند، معمولاً عاری از میکروب هستند. میکروب ها فقط در هنگام بیماری وارد این اندام ها می شوند.
باکتری ها در چرخه مواد
میکروارگانیسم ها به طور کلی و باکتری ها به طور خاص نقش بزرگی در چرخه های بیولوژیکی مهم مواد روی زمین بازی می کنند و دگرگونی های شیمیایی را انجام می دهند که برای گیاهان یا حیوانات کاملاً غیرقابل دسترسی است. مراحل مختلف چرخه عناصر توسط موجودات مختلف انجام می شود. وجود هر گروه منفرد از موجودات وابسته به تبدیل شیمیایی عناصر انجام شده توسط گروه های دیگر است.
چرخه نیتروژن
تبدیل چرخه ای ترکیبات نیتروژن دار نقش اصلی را در تامین اشکال لازم نیتروژن برای موجودات بیوسفر با نیازهای تغذیه ای متفاوت ایفا می کند. بیش از 90 درصد از تثبیت نیتروژن کل به دلیل فعالیت متابولیک باکتری های خاص است.
چرخه کربن
تبدیل بیولوژیکی کربن آلی به دی اکسید کربن، همراه با کاهش اکسیژن مولکولی، نیازمند فعالیت متابولیکی مشترک میکروارگانیسم های مختلف است. بسیاری از باکتری های هوازی اکسیداسیون کامل مواد آلی را انجام می دهند. در شرایط هوازی، ترکیبات آلی ابتدا با تخمیر تجزیه میشوند و در صورت وجود گیرندههای هیدروژن معدنی (نیترات، سولفات یا CO2)، محصولات نهایی آلی تخمیر با تنفس بیهوازی بیشتر اکسید میشوند.
چرخه گوگرد
گوگرد عمدتاً به شکل سولفات های محلول یا ترکیبات گوگرد آلی احیا شده در دسترس موجودات زنده است.
چرخه آهن
برخی از بدنه های آب شیرین حاوی غلظت بالایی از نمک های آهن کاهش یافته هستند. در چنین مکان هایی، یک میکرو فلور باکتریایی خاص ایجاد می شود - باکتری آهن، که آهن کاهش یافته را اکسید می کند. آنها در تشکیل سنگهای آهن باتلاقی و منابع آبی غنی از نمکهای آهن شرکت میکنند.
باکتری ها قدیمی ترین موجودات هستند که حدود 3.5 میلیارد سال پیش در آرکئن ظاهر شدند. برای حدود 2.5 میلیارد سال آنها بر زمین تسلط داشتند و زیست کره را تشکیل دادند و در تشکیل اتمسفر اکسیژن شرکت داشتند.
باکتری ها یکی از ساده ترین موجودات زنده (به جز ویروس ها) هستند. اعتقاد بر این است که آنها اولین موجوداتی هستند که روی زمین ظاهر شدند.
بر اساس سیستم جدید، سه حوزه متمایز می شوند: "باکتری ها" (یوباکتری ها)، "آرکئا" (آرکائوباکتری ها) و "ایزاگوا" (یوکاریوت ها). دامنه ها شامل فیلا، کلاس ها، راسته ها، خانواده ها، جنس ها و گونه ها هستند.
باکتری هاپروکاریوت هستند (در یک سلول پروکاریوتی، هسته به نام نوکلوئید دارای پوشش هسته ای، هسته و هیستون نیست و سیتوپلاسم حاوی اندامک های بسیار سازمان یافته نیست).
به دیواره ضخیم، گرم مثبت باکتری هاشامل: اشکال کروی یا کوکسی (استافیلوکوک، استرپتوکوک ها، پنوموکوک)؛ میله ای شکل، از جمله کورینه باکتری ها، مایکوباکتری ها و بیفیدوباکتری ها; اکتینومیست ها (باکتری های شاخه دار، رشته ای).
اشکال کروی یا کوکسی - باکتری های کروی با اندازه 0.5-1.0 میکرون؛ با توجه به آرایش نسبی سلول ها، میکروکوکس، دیپلوکوک، استرپتوکوک ها، تتراکوک ها، سارسینا و استافیلوکوک ها. میکروکوکس (یونانی) میکروس- کم اهمیت)
- سلول های جداگانه یا به شکل "بسته ها" واقع شده اند.
دیپلوکوکی (از یونانی.دیپلوها- کوکسی های دوتایی) یا جفتی که به صورت جفت چیده شده اند (پنوموکوک، گونوکوکمننگوکوک)، از آنجایی که سلول ها پس از تقسیم از هم جدا نمی شوند. پنوموکوکدر طرفین هم شکل نیزه ای دارد و گونوکوکو مننگوکوکشکل دانه های قهوه با سطح مقعر آنها رو به روی یکدیگر است. استرپتوکوک ها(از یونانی استرپتوز- زنجیره) - سلول هایی به شکل گرد یا کشیده که به دلیل تقسیم سلولی در همان صفحه زنجیره ای را تشکیل می دهند و ارتباط بین آنها را در محل تقسیم حفظ می کنند. Sarcins (از لات. سارسینا- دسته، عدل) به شکل "بسته هایی" از 8 یا بیشتر کوکسی چیده شده اند، زیرا آنها در طول تقسیم سلولی در سه صفحه متقابل عمود بر هم تشکیل می شوند. استافیلوکوک (از یونانی. استافیل- خوشه انگور) - کوکسی که به صورت خوشه انگور در نتیجه تقسیم در صفحات مختلف قرار دارد.
باکتری های میله ای شکلدر اندازه، شکل انتهای سلول و موقعیت نسبی سلول ها متفاوت است. طول سلول ها از 1.0 تا 8.0 میکرومتر، ضخامت - از 0.5 تا 2.0 میکرومتر متغیر است. میله ها می توانند به شکل منظم (اشریشیا کلی و غیره) و نامنظم (کورین باکتری و غیره) باشند، از جمله شکل های منشعب، به عنوان مثال در اکتینومیست ها. میله های کمی خمیده ویبریو (Vibrio cholerae) نامیده می شوند. بیشتر میله ای شکل باکتری هابه طور تصادفی قرار دارد، زیرا پس از تقسیم سلول ها از هم جدا می شوند. اگر پس از تقسیم، سلول ها توسط قطعات مشترک دیواره سلولی به هم متصل باقی بمانند و از هم جدا نشوند، در این صورت در زاویه ای نسبت به یکدیگر قرار می گیرند (کورین باکتریوم دیفتری) یا تشکیل یک زنجیره (باسیل سیاه زخم) می دهند.
برنج. 3.1. اسمیراز جانباشرشیاکلیواستافیلوکوکوس اورئوس.نقاشی گرم
در کتابچه راهنمای قدیمی باکتریولوژی سیستماتیک Bergey، باکتری ها بر اساس ویژگی های دیواره سلولی باکتری به 4 بخش تقسیم شدند: Gracilicutes - یوباکتری با دیواره سلولی نازک، گرم منفی. Firmicutes - یوباکتری با دیواره سلولی ضخیم، گرم مثبت؛ Tenericutes - یوباکتری بدون دیواره سلولی. مندوزیکوت ها آرکی باکتریایی با دیواره سلولی معیوب هستند. هر بخش بر اساس رنگ آمیزی گرم، شکل سلولی، نیاز به اکسیژن، تحرک، متابولیک و ویژگی های تغذیه ای به بخش ها یا گروه هایی تقسیم شد.
جدول 3.1. طبق ویرایش دوم (2001) کتابچه راهنمای Bergey، باکتری ها به دو حوزه تقسیم می شوند:باکتری ها»
و «
آرکیا»
|
دامنه "باکتری"(اوباکتری) |
دامنه "آرکیا"(باکتری ها) |
|
در دامنه «
باکتری ها»
باکتری های زیر را می توان جدا کرد: |
آرکی باکتری ها در دیواره سلولی خود پپتیدوگلیکان ندارند. آنها ریبوزوم های ویژه و RNA ریبوزومی (rRNA) دارند. اصطلاح "آرکائ باکتریا" در سال 1977 ظاهر شد. این یکی از اشکال باستانی زندگی است که پیشوند "arche" به معنای آن است. هیچ پاتوژن بیماری های عفونی در بین آنها وجود ندارد. |
|
اکثر باکتریهای گرم منفی بر اساس شباهتهای موجود در RNA ریبوزومی («پروتئوباکتریها» - از خدای یونانی پروتئوس، که اشکال مختلفی داشت) به شاخه پروتئوباکتریها دستهبندی میشوند. آنها از یک جد مشترک فتوسنتزی تکامل یافته اند. |
|
به دامنه "باکتری ها"شامل 22 نوع است که موارد زیر دارای اهمیت پزشکی هستند (به گفته برگی، 2001):
تایپ کنیدکه درIV. Deinococcus-ThermusکلاسI. Deinococci
راسته I. خانواده Deinococcales I. Deinococcaceae جنس1. دینوکوکوس
تایپ کنیدکه درXII. پروتئوباکتری ها
کلاسI. آلفاپروتئوباکتریسفارش II. خانواده Rickettsiales I. Rickettsiaceae جنس I. Rickettsia جنس I. Orientia جنس III. خانواده Wolbachia I. Ehrlichiaceae (b genera) جنس I. Ehrlichia جنس I. Aegyptianella خانواده III. Holosporaceae (8 جنس) راسته VI. خانواده Rhizobiales I. Bartonellaceae جنس I. Bartonella خانواده III. Brucellaceae جنس I. Brucella
کلاسII. بتاپروتئوباکتری هاراسته I. خانواده Burkholderiales I. Burkholderiaceae جنس I. خانواده Burkholderia IV. Alcaligenaceae جنس I. Alcaligenes جنس III. Bordetella ردیف IV. خانواده Neisseriales I. Neisseriaceae جنس I. Neisseria جنس VI.Eikenella جنس IX.Kingella راسته V. Nitrozomonadales خانواده II. Spirillaceae جنس I. Spirillum
کلاسIII. گاماپروتئوباکتری هاسفارش V. Thiotrichales خانواده III. Francisellaceae جنس I. Francisella Order VI. خانواده Legionellales I. Legionellaceae جنس I. Legionella Family II. Coxiellaceae جنس I. Coxiella راسته IX. خانواده Pseudomonadales I. Pseudomonadaceae جنس I. Pseudomonas خانواده I. Moraxellaceae جنس I. Moraxella جنس I. Acinetobacter ردیف XI. Vibrionales خانواده I. Vibrionaceae جنس I. Vibrio Order XII. خانواده Aeromonadales I. Aeromonadaceae جنس I. Aeromonas راسته XIII. خانواده انتروباکتریال ها I. انتروباکتریاسه جنس I. انتروباکتر جنس هشتم. Calymmatobacterium جنس X. سیتروباکتر جنس XI. ادواردزیلا جنس دوازدهم. Erwinia Genus XIII Escherichia Genus XV. هافنیا
جنس شانزدهم. کلبسیلا جنس XVII. Kluyvera جنس XXI. مورگانلا جنس XXVI. Plesiomonas جنس XXVIII. جنس پروتئوس XXIX. Providencia RodXXXY. سالمونلا جنس XXXIII. Serratia جنس XXXIV. Shigella Genus XL. یرسینیا سفارش IV. خانواده Pasteurellales I. Pasteurellaceae جنس I. Pasteurella جنس I. Actinobacillus جنس III. هموفیلوس
کلاسIV. دلتاپروتئوباکتری هاسفارش II. Desulfovibrionales خانواده I. Desulfovibrionaceae جنس II. بیلوفیلا
کلاسV. اپسیلون پروتئوباکتریراسته I. Campylobacteriales خانواده I. Campylobacteriaceae جنس I. Campylobacter Family II. Helicobacteriaceae جنس I. Helicobacter جنس Il.Wolinella
تایپ کنیدکه درسیزدهم. فیرمیکوت ها (اصلیمسیرگرم مثبت)
کلاسمن.
کلستریدیاراسته I. خانواده Clostridiales I. Clostridiaiaceae جنس I. Clostridium جنس IX. خانواده سارسینا III. Peptostreptococcaceae جنس I. Peptostreptococcus خانواده IV. Eubacteriaceae جنس I. Eubacterium خانواده V. Peptococcaceae جنس I. Peptococcus خانواده VII. Acidaminococcaceae جنس چهاردهم. ویلونلا کلاسII. مولیکوت هاراسته I. خانواده Mycoplasmatales I. Mycoplasmataceae جنس I. Mycoplasma جنس IV. اورهاپلاسما کلاسIII. باسیلراسته I. خانواده Bacilalles I. Bacillaceae جنس I. خانواده باسیلوس I. Planococcaceae جنس I. Planococcus جنس IV. Sporosarcina خانواده IV. Listeriaceae جنس I. Listeria خانواده V. Staphylococcaceae جنس I. Staphylococcus جنس II. Gemella Order I. Lactobacillus خانواده I. Lactobacillaceae جنس I. Lactobacillus جنس III. Pediococcus Family II. Aegococcusae جنس I. Aerococcus خانواده IV. Enterococcaceae جنس I. Enterococcus
خانواده V. Leuconostocaceae جنس I. Leuconostoc خانواده VI. Streptococcaceae جنس I. استرپتوکوکوس جنس II. لاکتوکوکوس تایپ کنیدکه درچهاردهم اکتینوباکتری هاکلاسI. اکتینوباکتری هازیر کلاس V. Actinobacteridae راسته I. Actinomycetales زیررده V. Actinomycineae خانواده I. Actinomycetaceae جنس I. Actinomyces جنس I. جنس Actinobacilum III. آرکانوداکتریومجنس IV. Mobiluncus فرعی VI. خانواده Micrococcineae I. Micrococcaceae جنس I. Micrococcus جنس VI. Rothia جنس VII. استوماتوکوکوس زیررده VII. خانواده Corynebacterineae I. Corynebacteriaceae جنس I. Corynebacterium خانواده I. Mycobacteriaceae جنس IV. مایکوباکتریوم خانواده V. Nocardiaceae جنس I. نوکاردیاجنس I. رودوکوکوس فرعی VII. خانواده Propionibacterineae I. Propionibacteriaceaeجنس I. پروپیونی باکتریخانواده I. جنس Nocardiaceae I. Nocardioidesسفارش II. بیفیدوباکتریال ها
خانواده I. Bifidobacteriaceae جنس I. Bifidobacterium جنس III. گاردنرلا تایپ کنیدکه درشانزدهم کلامیدیاکلاسI. کلامیدیاراسته I. خانواده Chlamydiales I. Chlamydiaceae جنس I. Chlamydia جنس II. کلامیدوفیلا تایپ کنیدکه درXVII. اسپیروکت هاکلاسI. اسپیروکت هاراسته I. خانواده Spirochaetales I. Spirochaetaceae جنس I. Spirochaeta جنس I. Borrelia جنس IX. خانواده ترپونما III. Leptospiraceae جنس I. Leptospira تایپ کنیدکه درXX. باکتریوئیدهاکلاسI. باکتریوئیدهاراسته I. خانواده Bacteroidales I. Bacteroidaceae جنس I. Bacteroides خانواده III. Porphyromonadaceae جنس I. Porphyromonas خانواده IV. Prevotellaceae جنس I. Prevotella کلاسII. فلاووباکتری هاراسته I. Flavobacteriales خانواده I. Flavobacteriaceae جنس I. Flavobacterium

*محل اسپور: 1 - مرکزی، 2 - زیر ترمینال، 3 - ترمینال.
برنج. 3.2.
کلامیدیامتعلق به باکتری های گرم منفی کوکوئید داخل سلولی اجباری (گاهی اوقات متغیر گرم) است. آنها فقط در سلول های زنده تولید مثل می کنند. در خارج از سلول ها، کلامیدیا کروی شکل (0.3 میکرومتر)، از نظر متابولیک غیرفعال هستند و اجسام ابتدایی نامیده می شوند. دیواره سلولی اجسام ابتدایی حاوی یک پروتئین غشای خارجی اصلی و یک پروتئین حاوی مقدار زیادی سیستئین است. اجسام ابتدایی با تشکیل یک واکوئل درون سلولی با اندوسیتوز وارد سلول اپیتلیال می شوند. در داخل سلول ها، آنها بزرگ شده و به اجسام مشبک در حال تقسیم تبدیل می شوند و خوشه هایی در واکوئل ها (شامل ها) تشکیل می دهند. اجسام ابتدایی از اجسام مشبک تشکیل می شوند که در اثر اگزوسیتوز یا لیز سلولی از سلول خارج می شوند. اجسام ابتدایی آزاد شده از سلول وارد چرخه جدیدی می شوند و سلول های دیگر را آلوده می کنند. کلامیدیا در انسان باعث آسیب به چشم ها، دستگاه ادراری تناسلی، ریه ها و غیره می شود.
مایکوپلاسما -باکتری های کوچک (0.15-1.0 میکرومتر)، که توسط یک غشای سیتوپلاسمی احاطه شده و بدون دیواره سلولی است. به دلیل عدم وجود دیواره سلولی، مایکوپلاسماها از نظر اسمزی حساس هستند. آنها اشکال مختلفی دارند: کوکوئید، رشته ای، فلاسکی شکل. شبیه به اشکال L (شکل 3.6). این اشکال در میکروسکوپ فاز کنتراست کشت خالص مایکوپلاسما قابل مشاهده است. مایکوپلاسماهای بیماری زا باعث عفونت های مزمن - مایکوپلاسموز می شوند.
اکتینومیست ها -باکتری های گرم مثبت شاخه دار، رشته ای یا میله ای شکل. نام آن (از یونانی. عمل می کند- اشعه، مایکز- قارچ) آنها را به دلیل تشکیل در بافت های آسیب دیده دروسن دریافت کردند - گرانول های نخ های محکم در هم تنیده به شکل پرتوهایی که از مرکز گسترش یافته و به ضخامت های فلاسکی شکل ختم می شود. اکتینومیست ها می توانند با تکه تکه شدن میسلیوم به سلول هایی شبیه باکتری های میله ای شکل و کوکوئید تقسیم شوند. روی هیف های هوایی اکتینومیست ها، هاگ هایی تشکیل می شوند که برای تولید مثل استفاده می شوند. اسپورهای اکتینومیست معمولاً در برابر حرارت مقاوم نیستند.

محل اتصال (مشابه بلفاروپلاست)
برنج. 3.3.الگوی پراش الکترونی یک قطعه سلولی ترپونما پالیدوم (کنتراست منفی). به گفته N. M. Ovchinnikov، V. V. Delectorsky
یک شاخه فیلوژنتیک مشترک با اکتینومیست ها توسط اکتینومیست های به اصطلاح نوکاردیفرم (نوکاردیوفورم) تشکیل می شود - گروهی از باکتری های میله ای شکل و نامنظم. نمایندگان فردی آنها فرم های شاخه ای را تشکیل می دهند. اینها شامل باکتری های این جنس است کورینه باکتریوم,
مایکوباکتریوم,
نوکاردیااکتینومیست های شبه نوکاردی با وجود قندهای آرابینوز، گالاکتوز و همچنین اسیدهای مایکولیک و مقادیر زیادی اسیدهای چرب در دیواره سلولی متمایز می شوند. اسیدهای مایکولیک و لیپیدهای دیواره سلولی مقاومت اسیدی باکتری ها، به ویژه مایکوباکتریوم توبرکلوزیس و جذام را تعیین می کنند (در صورت رنگ آمیزی طبق Ziehl-Neelsen، آنها قرمز هستند و باکتری ها و عناصر بافتی غیر مقاوم به اسید، خلط آبی هستند).
اشکال پیچ خورده باکتری های مارپیچی شکل هستند، به عنوان مثال اسپیرلا، که ظاهر سلول های پیچ خورده چوب پنبه ای شکل دارند. اسپیریلای بیماری زا شامل عامل ایجاد کننده سودوکو (بیماری نیش موش) است. پیچیدگی ها نیز شامل کمپیلوباکتر، هلیکوباکتر هستند که دارای انحناهایی مانند بال مرغ دریایی در حال پرواز هستند. باکتری هایی مانند اسپیروکت ها نیز به آنها نزدیک هستند. اسپیروکت ها باکتری های نازک، دراز و پیچ خورده (مارپیچی شکل) هستند که به دلیل تغییرات خمشی در سلول ها، از نظر تحرک با اسپیریلوم تفاوت دارند. اسپیروکت ها یک دیواره سلولی غشایی خارجی دارند که یک استوانه پروتوپلاسمی را با یک غشای سیتوپلاسمی احاطه کرده است. در زیر غشای خارجی دیواره سلولی (در پری پلاسم) فیبریل های پری پلاسمیک (تاژک) وجود دارد که گویی به دور استوانه پروتوپلاسمی اسپیروکت می چرخند، به آن شکل مارپیچ می دهند (فرش های اولیه اسپیروکت). فیبریل ها به انتهای سلول متصل می شوند (شکل 3.3) و به سمت یکدیگر هدایت می شوند. انتهای دیگر فیبریل ها آزاد است. تعداد و آرایش فیبریل ها در بین گونه ها متفاوت است. فیبریل ها در حرکت اسپیروکت ها نقش دارند و به سلول ها حرکت چرخشی، خمشی و انتقالی می دهند. در این حالت، اسپیروکت ها حلقه ها، فرها و خم هایی را تشکیل می دهند که به آنها فرهای ثانویه می گویند.
اسپیروکت هارنگ ها را خوب قبول نکنید آنها با استفاده از روش رومانوفسکی-گیمسا یا آبکاری نقره رنگآمیزی میشوند و به صورت زنده با استفاده از میکروسکوپ فاز کنتراست یا میدان تاریک مورد بررسی قرار میگیرند. اسپیروکت ها توسط 3 جنس که برای انسان بیماری زا هستند نشان داده می شوند: ترپونما, بورلیا, لپتوسپیرا.
ترپونما(جنس Treponema) دارای ظاهری نخ های نازک و پیچ خورده با چوب پنبه با 8 تا 12 فر کوچک یکنواخت است. فیبریل ها در اطراف پروتوپلاست ترپونما قرار دارند. نمایندگان بیماری زا T. پالیدوم- عامل ایجاد کننده سیفلیس، تی. همیشگی- عامل بیماری استوایی یاوز.
بورلیا(جنس Borrelia) بلندتر، دارای 3-8 حلقه بزرگ و 8-20 فیبریل است. اینها شامل عامل ایجاد کننده تب عود کننده است (ب. عود کننده) و پاتوژن های بیماری لایم (V. بورگدورفریو غیره.).
لپتوسپیرا(جنس Leptospira) دارای فرهای کم عمق و مکرر هستند - به شکل یک طناب پیچ خورده. انتهای این اسپیروکت ها مانند قلاب هایی خمیده هستند که در انتهای آن ها ضخیم می شوند. با تشکیل فرهای ثانویه، آنها ظاهر حروف "S" یا "C" را به خود می گیرند. دارای 2 رشته محوری نماینده بیماری زا L
